
Pteroptyx bearni Olivier, 1966
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3653.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:72A07BC6-AEB0-4EBC-AFA8-F5871065680F |
|
DOI |
https://doi.org/10.5281/zenodo.5266897 |
|
persistent identifier |
https://treatment.plazi.org/id/E46FF216-5E65-C527-FF6F-BD9CFF09EECF |
|
treatment provided by |
Felipe |
|
scientific name |
Pteroptyx bearni Olivier |
| status |
|
Pteroptyx bearni Olivier View in CoL
[ Figs 191–201, 204, 205 View FIGURES 191–199 View FIGURES 200–207 ]
Pteroptyx Bearni Olivier View in CoL in Olivier and Pic, 1909:139; 1910:47.
Pteroptyx bearni Olivier. McDermott 1966:117 View in CoL . Ballantyne & McLean 1970:241 (Partim). Ballantyne 2001:62 (Partim). Ohba &Wong 2012:.
Pteroptyx similis Ballantyne. Ballantyne 2001:62 View in CoL . Mahadimenakbar, Hairul & Mazidi (2007):1. (New synonymy).
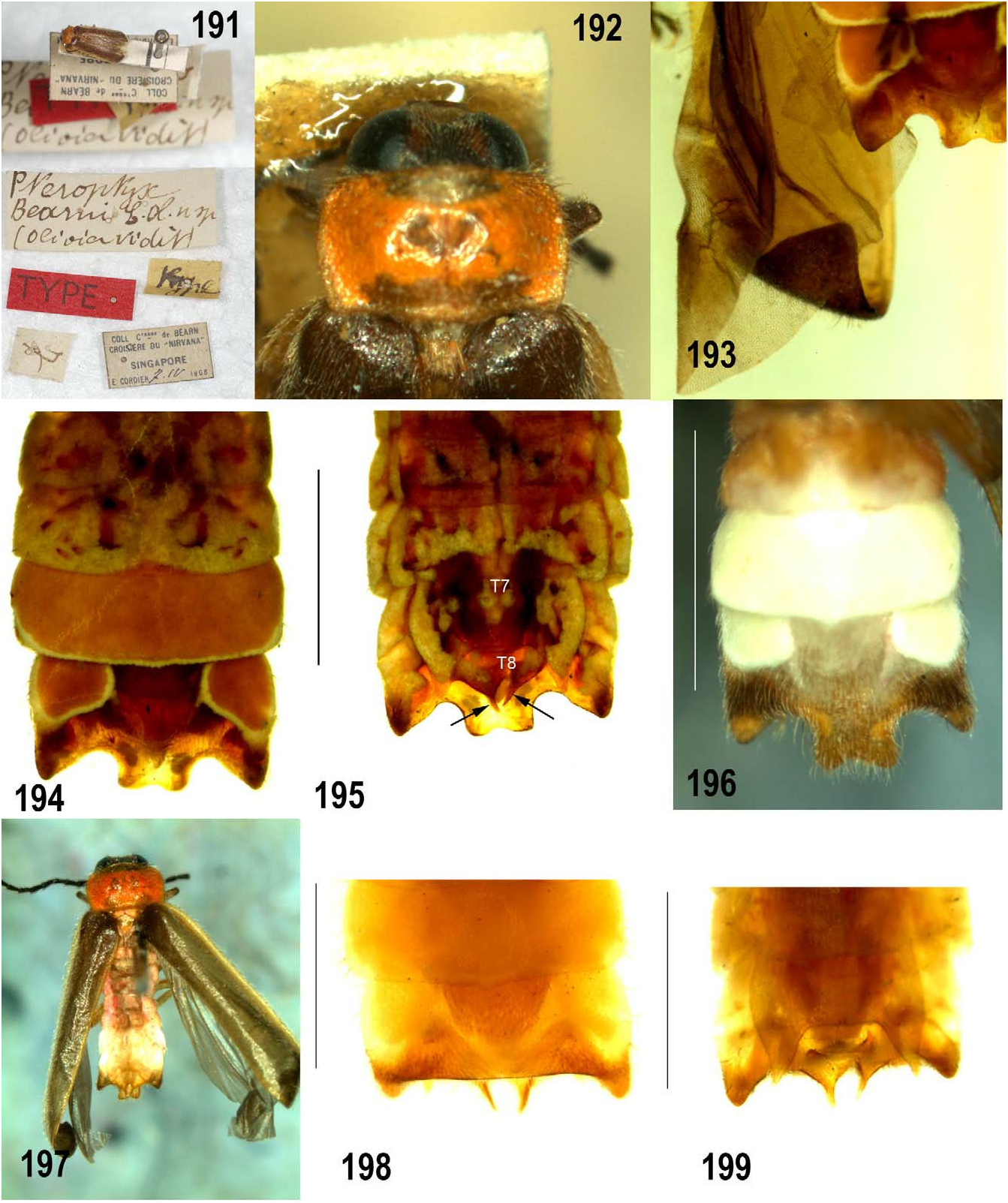
Lectotype. Male. SINGAPORE: Labelled 1. Handwritten: Pteroptyx bearni E. Ol. View in CoL n. sp. (Olivier vidit) 2. Red printed label: TYPE. 3. Handwritten: type. 4. Symbol. 5. Printed: Coll. Ctesse de bearn Croisvere du “Nirvana” Singapore E. Cordier 7.iv.1908 ( MNHN).
Other specimens examined. MALAYSIA: Kalabakan River , 2.vii.1972, I. Polunin, 10 males, 7 females ( ANIC). Pingan Pingan, 20.vi.2009, Vito, male ( FRIM) .
Diagnosis. Belonging to a group of four species of Pteroptyx s. str. in which T8 has elongate lobes beside the median posterior emargination; most obviously distinguished from asymmetria and tener by the shape of the terminal abdominal tergites (in asymmetria T7 is not medially emarginated, and T8 is asymmetrical; in tener T7 is shallowly emarginated and slightly sinuous with posterolateral corners narrowed, and T8 is symmetrical); similar to P. decolor which is not well characterised in collections, differing in the dorsal colour pattern ( P. decolor is entirely pale coloured dorsally).
Lectotype male. 5.5 mm long; Colour ( Figs 191, 192, 194, 195 View FIGURES 191–199 ): pronotum ( Fig. 192 View FIGURES 191–199 ) orange yellow (fat body retracted along both anterior and posterior margins); MS and MN pale brownish; elytra semitransparent, light brown with narrowly pale lateral margin not extending around apex; head, antennae and palpi very dark brown; ventral thorax and legs brownish orange except for dark brown tibiae and tarsi 1, dark brown tarsi 2 and dark brown 4 th and 5 th tarsomeres of legs 3; ventral thoracic colour (especially that of metathorax) partly obscured by dark dehydrated thoracic muscles visible beneath cuticle; basal abdominal ventrites whitish-yellow (underlying fat body visible beneath semitransparent cuticle contributes to colour); LO ( Fig. 194 View FIGURES 191–199 ) in V6, 7 yellowish white with very white margins; remainder of V7 orange; abdominal tergites ( Fig. 195 View FIGURES 191–199 ) orange, cuticle semitransparent and colour obscured by underlying white fat body; dorsal aspect of apices of PLP narrowly brown ( Fig. 195 View FIGURES 191–199 ). Pronotum ( Fig. 192 View FIGURES 191–199 ): 0.9 mm long; 1.5 mm wide; W/L=1.6; median anterior margin rounded scarcely projecting beyond obtusely rounded anterolateral corners; lateral margins slightly convex sided (i.e. B>A, C); median posterior margin rounded, projecting beyond angulate obtuse posterolateral corners, and separated from them by shallow emarginations; depressed area visible in median area of disc probably post-mortem artefact. Elytra: parallel sided; deflexed apex truncate with inner corner rounded ( Fig. 193 View FIGURES 191–199 ). Head: GHW 1.1 mm; SIW 0.3mm; ASD subequal to ASW; apical labial palpomere slightly longer than wide and ½ as long as apical maxillary palpomere. Antennae incomplete; FS1 elongate slender as long as scape. Legs: MFC sparse, may have up to 5 teeth (teeth appear to be missing).Abdomen ( Figs 194, 195 View FIGURES 191–199 ): posterior margin of V7 with broad MPP which is apically shallowly emarginated, and bearing tiny hooks on the dorsal surface of the posterolateral corners; MPP projecting posteriorly just beyond narrowed apices of PLP; LOs in V7 separated in middle by their width and occupying less than half the area of V7; posterior margin of T7 deeply and widely emarginated with posterolateral corners narrowing and apically acute; lateral margins of visible portion of T8 rounded, median posterior margin emarginated, emargination bounded laterally by narrowed apically pointed projections. Aedeagal complex not dissected from this unique specimen. Based on Ballantyne and McLean (1970), Ballantyne (2001), the aedeagal index (B/A) for bearni is 0.85 and 0.6 for P. tener .
Female ( Figs 200, 201, 204, 205 View FIGURES 200–207 ): (Kalabakan River specimens). 4.8–6.3 mm long. Colour: as for male except for pale LO in V6 only, and pale yellow slightly transparent V7, 8; no pink colouration in pronotum or fat body discerned. Bursa: anterior plates appearing indented along margins, probably postmortem effect ( Figs 200, 201 View FIGURES 200–207 ).
Discussion. Ballantyne (2001) distinguished P. similis from both P. decolor and P. tener , but not from P. bearni which had not been adequately characterised in Ballantyne and McLean (1970). Tergite 8 was not dissected and Ballantyne and McLean’s Figure 6d View FIGURE 6 shows T7 outline consistent with that of bearni as described here, but T8 is depicted without lobes to either side of the median emargination. Ballantyne (2001; Fig. 18 View FIGURES 18–25 ) depicted a male from Paya Paloh apparently lacking these lobes on T8 and its identity has not been confirmed.
Ohba and Wong (2012) indicated incorrectly that the morphological features of this and P. tener were very similar except for the orange colour in the pronotum.
In the field the distinctive pinkish –orange pronotum and darker elytra enable quick and usually accurate identification of P. bearni (Wong pers com.), especially if light patterns are observed (e.g. Fig. 197 View FIGURES 191–199 ). Ballantyne (in prep.) observed difficulties interpreting pronotal colour in ethanol preserved specimens, where the pink colour may have leached out. Its occurrence may be variable, since pinned specimens from the same locality also vary in pronotal colour. The pink colour in the fat body is usually found elsewhere apart from the pronotum, and can be seen especially in the dorsal abdomen, where the cuticle is paler and semitransparent (e.g. Fig. 197 View FIGURES 191–199 ). Specimens of P asymmetria from Perak and Selangor (FRIM) also have pinkish colour in the pronotum (Ballantyne obs.).
In one Kalabakan River specimen the posterior margin of V7 is arched dorsally such that MPP not visible and projections of T8 are visible from beneath ( Figs 198, 199 View FIGURES 191–199 ).
P. bearni is non-synchronous, and is usually found downstream from the synchronously flashing P. tener in the more saline waters, but occasionally, probably due to tidal inflows, their habitats may overlap ( Ohba and Wong 2012).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pteroptyx bearni Olivier
| Ballantyne, Lesley A. & Lambkin, Christine L. 2013 |
Pteroptyx similis
| Mahadimenakbar, M. D. & Hairul, H. M. & Mazidi, A. 2007: 1 |
Pteroptyx bearni Olivier. McDermott 1966:117
| Ballantyne, L. A. & McLean, M. R. 1970: 241 |
| McDermott, F. A. 1966: 117 |
Pteroptyx Bearni Olivier
| Olivier, E. & Pic, M. 1909: 139 |