
Palaeoneura Waterhouse 1915
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1455.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C91CD45A-6019-4070-BF32-61E17543C5D0 |
|
persistent identifier |
https://treatment.plazi.org/id/E063C61C-FFE3-FFEA-FF0D-F9044A136059 |
|
treatment provided by |
Felipe |
|
scientific name |
Palaeoneura Waterhouse 1915 |
| status |
|
Palaeoneura Waterhouse 1915 View in CoL View at ENA
(Figs 73–76, 78–83)
Palaeoneura Waterhouse 1915: 537–538 View in CoL ; Soyka 1956: 107; Annecke & Doutt 1961: 30–31. Type species: P. interrupta Waterhouse View in CoL , designated by Gahan & Fagan 1923: 103.
Chaetomymar Ogloblin 1946: 277 View in CoL ; Annecke & Doutt 1961: 34; Huber 2003: 78–81 (taxonomic history, diagnosis, key to species, etc.). Type species: C. kusnezovi Ogloblin View in CoL , by original designation. Syn. n.
Acanthomymar Subba Rao 1970: 667–668 View in CoL ; Hayat & Anis 1999: 307; Huber 2003: 80 (discussion, synonymy under Polynema View in CoL ). Type species: A. nigrum Subba Rao , by original designation. Syn. n. (from previous synonymy under Polynema Haliday View in CoL by Huber 2003: 80).
Redescription
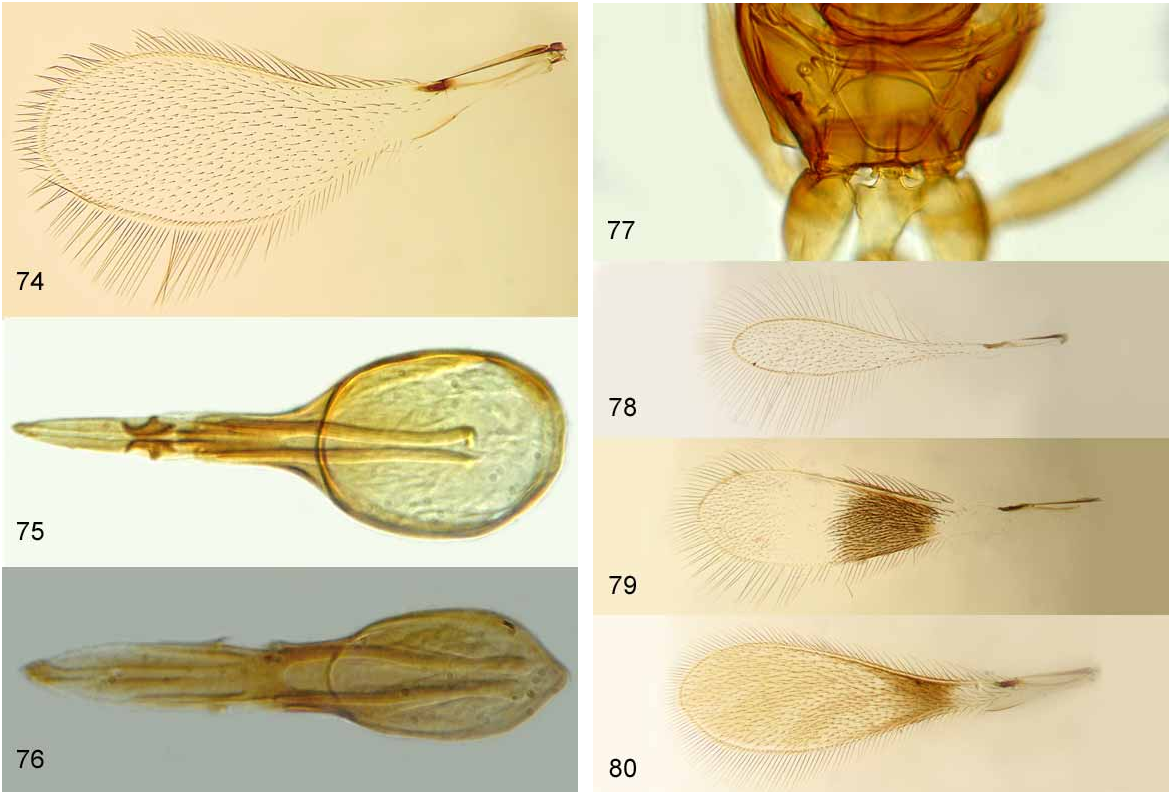
Body color yellow to dark brown or black. Face usually smooth, without a pit next to each torulus, but sometimes with a small medial groove extending from transverse trabecula down to about level of toruli and fine sculpture perpendicular to and next to (from both sides) this groove ( Fig. 81 View FIGURES 81–83 ). Mandible tridentate. Female funicle 6-segmented, all segments usually much longer than wide, longitudinal sensilla sometimes present on F6 or, rarely, on F5 and F6; clava entire, with 7 to 9 longitudinal sensilla. Propleura abutting each other anteriorly along midline, the prosternum thus closed anteriorly (Fig. 73); mesonotum usually smooth; mesoscutum with distinct notauli; scutellar sensilla usually somewhat in middle of scutellum but sometimes at its anterior margin; propodeum smooth, without carinae. Forewing characteristic, often narrow (Fig. 78), but sometimes broad ( Fig. 74 View FIGURES 74–76 ), and with at least a slight narrowing just beyond apex of venation; marginal vein relatively long and usually with 2 dorsal macrochaetae; chaetotaxy of forewing blade variable, with or without conspicuous dark bands or spots and often with a slight but conspicuous darkening extending as a line along anterior margin and not reaching wing apex distally or apex of venation basally (Figs 80, 83). Petiole longer than wide, attached posteriorly to gastral tergum; male genitalia of characteristic shape ( Figs 75, 76 View FIGURES 74–76 ).
FIGURES 77–80. Mymaridae . 77. Polynema (Polynema) editha (female, Australia), propodeum and petiole. 78–80. Palaeoneura spp. 78. P. silvifilia (male, Australia), forewing. 79. P. sappho (female, Australia), forewing. 80. P. sp. (male, Tahiti, French Polynesia), forewing.
Diagnosis
Superficially, Palaeoneura is most likely to be confused with the nominate subgenus of Polynema . But P. ( Polynema ) has the propleura not abutting anteriorly, with the prosternum thus open anteriorly ( Fig. 124 View FIGURES 122–124 ), a short marginal vein, the forewing blade usually wider just beyond apex of venation than at marginal vein (Fig. 125), and the propodeum usually with a medial carina, either incomplete (Fig. 77) or complete.
Comments
Both Soyka (1956) and Annecke & Doutt (1961) erroneously indicated Palaeoneura turneri Waterhouse as the type species of Palaeoneura . It indeed would had been a better choice as type species because it was described by Waterhouse (1915) on the same page but prior to P. interrupta and the darkening of the forewing membrane is more conspicuous in P. turneri . But because the original generic description of Palaeoneura is so vague that it can be applied to both of the two very distinctive genera that comprise Waterhouse's Palaeoneura , and to avoid any other unnecessary confusion, we accept Gahan & Fagan's (1923) designation of P. interrupta as the type species of Palaeoneura and describe below a new genus, Boccacciomymar , based on P. turneri .
Biology
Host associations and other biological information are unknown for the majority of the members of Palaeoneura but the species formerly placed in Chaetomymar are egg parasitoids of various Cicadellidae , as listed by Huber (2003).
Distribution and taxonomic remarks
Afrotropical, Australasian (including New Zealand and Oceania), Oriental, eastern and southern (Arabian Peninsula) Palaearctic; occasional in the Neotropics (several undescribed species). Palaeoneura is extremely common in Australia, where it comprises most of the 50 species classified before in Polynema . In fact, Polynema (Polynema) , as defined by Triapitsyn & Fidalgo (2006), is quite rare in Australia (and also New Zealand); we have seen Australian specimens belonging to several species, three of which are P. draperi Girault , P.editha Girault and P.frater Girault. The native species of Polynema (Polynema) from Australia and New Zealand seem to belong to a distinct species group with a peculiar petiole with dorsolateral membranous "wings" anteriorly (Fig. 77) and also usually just one dorsal seta on the marginal vein (although sometimes their number may vary from one to two even in the same specimen). Polynema mendeli Girault is a member of Polynema (Dorypolynema) Hayat & Anis ( Hayat & Anis 1999) ; P. aequum Girault was transferred to Himopolynema Taguchi ( Triapitsyn & Berezovskiy 2002). Richteria ara (Girault) (= Polynema blackbourni Girault , syn. n. and P. helena Girault , syn. n.) is transferred to Richteria Girault , comb. n. from Polynema ; different forewing color patterns in this species are due to sexual dimorphism (J.T. Huber, personal communication; also based on the two specimens from the same collection event, a female and a male, with the following data: Australia, Queensland, Brisbane Forest Park, Mt. Glorious Rd. W of Lomandra Junction, 27°23.57’S, 152°54.68’E, 5–6.xii.2002, J. George, J. Munro, A. Owen [UCRC]). Agalmopolynema elatum (Girault) , A. glabricorpus (Girault) , A. grotiusi (Girault) , A. lodgei (Girault) , and A. wallacei (Girault) are transferred to Agalmopolynema , all comb. n. from Polynema . These are all good species, distinguished from each other by female morphology (we have identified recently collected specimens belonging to all of them [in ANIC, CNCI, and UCRC]). The Australian species of Agalmopolynema have two macrochaetae on the marginal vein (a more primitive character state, as in many Oriental species of Acmopolynema ) whereas the South American species of Agalmopolynema all have one macrochaeta. The remaining species of Girault's Australian Polynema are transferred below either to Palaeoneura or Boccacciomymar gen. n.
Checklist of Palaeoneura species
australiense (Girault) ( Polynema ), comb. n.
bagicha (Narayanan, Subba Rao & Kaur) ( Polynema , from Chaetomymar ), comb. n.
breviscapus (Girault) ( Polynema ), comb. n.
darwini (Girault) (Polynema) , comb. n.
dei (Girault) ( Polynema , from Chaetomymar ), comb. n.
devriesi (Girault) ( Polynema ), comb. n.
elisabethae (Ferrière) ( Polynema ; from Chaetomymar ), comb. n.
filia (Girault) ( Polynema filius ), comb. n.
franklini (Girault) ( Polynema ), comb. n.
goldsmithi (Girault) ( Polynema silvifilia goldsmithi ), comb. n. & stat. n.
gracile (Prinsloo) ( Chaetomymar ), comb. n.
haeckeli (Girault) ( Polynema ), comb. n.
hegeli (Girault) (Polynema) , comb. n.
hishimoni (Taguchi) ( Chaetomymar ), comb. n.
indopeninsularis (Mani & Saraswat) ( Polynema , from Chaetomymar ), comb. n.
joulei (Girault) ( Polynema ), comb. n.
kusnezovi (Ogloblin) (Chaetomymar) , comb. n.
lepida (Annecke & Doutt) ( Chaetomymar lepidum ), comb. n.
mendeleefi (Girault) ( Polynema ), comb. n.
nigra (Subba Rao) ( Acanthomymar nigrum ; Polynema nigrum , from Polynema ), comb. n. nigrithorax (Girault) ( Polynema poincarei nigrithorax ), comb. n. & stat. n.
nordaui (Girault) (Polynema) , comb. n.
notabilissima (Girault) ( Polynema notabilissimum ), comb. n.
pax (Girault) ( Polynema ), comb. n.
poincarei (Girault) (Polynema) , comb. n.
romanesi (Girault) ( Polynema ), comb. n.
rousseaui (Girault) ( Polynema ), comb. n.
sappho (Girault) (Polynema) , comb. n.
shakespearei (Girault) ( Polynema ), comb. n.
sieboldi (Girault) ( Polynema ), comb. n.
signa (Girault) ( Polynema signum ), comb. n.
silvae (Girault) ( Polynema ), comb. n.
silvifilia (Girault) (Polynema) , comb. n.
sophoniae (Huber) ( Chaetomymar ), comb. n.
speciosissima (Girault) ( Polynema speciosissimum ), comb. n.
spenceri (Girault) (Polynema) , comb. n.
stupenda (Girault) ( Polynema stupendum ), comb. n.
sublesta (Girault) ( Polynema sublestum ), comb. n.
thoreauini (Girault) ( Polynema ), comb. n.
unimaculata (Hayat & Anis) ( Acmopolynema unimaculatum ), comb. n.
virgilii (Girault) (Polynema) , comb. n.
zangwilli (Girault) ( Polynema ), comb. n.
zolai (Girault) (Polynema) , comb. n.
Comments
The nomenclatural changes proposed here are based on study of the types of Girault's Australian species of Polynema during the senior author's trip to Australia in January 1999, subsequent loans of type material from QMBA, New's (1976) review (particularly his illustrations), and also the study of more than a thousand specimens of Polynema -like mymarids from ANIC, CNCI, QMBA and UCRC; the vast majority of these specimens turned out to be Palaeoneura . The generic placement of P. virgilii is unclear because its type is lost ( New 1976; Dahms 1986). According to the very brief original description by Girault (1938), it has F2 and F3 equal in length. Because F2 is notably longer than F3 (and the longest funicle segment) in all known Australian species of Polynema (Polynema) , it is very unlikely that P. virgilii belongs there, so we transfer it to Palaeoneura as that is most likely its correct placement. The placement of P. hegeli , P. poincarei and P. nigrithorax in Palaeoneura is also tentative because some of their features, such as placement of the toruli very close to the transverse trabecula, presence of relatively large, apically blunt supraorbital setae on the head, and structure of the male genitalia suggest their resemblance to the New World genus Neomymar Crawford , whereas the shape of the forewing and some other features suggest that these species rather might be just unusual members of Palaeoneura . Until molecular and additional morphological evidence becomes available, we prefer taking a conservative approach, at least temporarily classifying them in Palaeoneura . They clearly do not belong in Polynema . When all Old World Neomymar -like mymarids are reviewed (they occur in the Oriental region, mainly in Indonesia and Papua New Guinea, and also in Madagascar), these three species may be placed in a new genus. Placement of P. notabilissimum , P. spenceri , and P.zolai in Palaeoneura is also tentative; the type specimens of the latter two species need to be remounted properly (after dissolving the old Canada balsam and subsequent clearing) in order to determine their correct generic placement. It is quite possible, for instance, that P. zolai will turn out to be an unusual Acmopolynema but to show that (or otherwise), a dorsoventral remount of the holotype is needed (to be able to see the propodeum in dorsal view).
Once we determined that most of A.A. Girault's Australian species of Polynema belong in Waterhouse's Palaeoneura , the necessity for the proposed synonymies of Chaetomymar Ogloblin and Acanthomymar Subba Rao became obvious. They seem to represent at most distinct species groups within the very large and diverse genus Palaeoneura . One group within Palaeoneura is comprised of the species formerly placed in Chaetomymar , revised by Huber (2003), P. nordaui , and also many undescribed species; all these have mostly yellow to light brown bodies (although at least one species from Australia has a brown body in females), a very long female antenna with a dark brown clava, and a very narrow forewing with or without various dark markings. The main distinguishing feature of this group is the very long, strong, axillar seta extending at least to the level of the posterior margin of the scutellum; this seta is particularly wide basally (in lateral view). The second species group is a loose conglomeration of the species resembling P. nigra and many similarly Acanthomymar -looking undescribed species from Africa and Madagascar; this group also includes several Oriental species (such as P. unimaculata ), a few undescribed Neotropical species, and many of the Australian species (such as P. silvifilia ). It is defined mainly by the very long female antenna, a narrow forewing, usually with the characteristic arrangement of setae just distal to the tip of venation (as in Fig. 78) and a moderately long and strong, usually blunt axillar seta that usually does not reach (or barely reaches) the posterior margin of the scutellum; the body color and presence or absence of markings on the forewing vary a lot in this species group. Among the African and Australasian Acanthomymar -like species of Palaeoneura , the notauli can be very narrow (as in P. nigra ), moderately broad (as in some undescribed species from Kenya and Madagascar), or broad (as in P. silvifilia and P. unimaculata ). The third species group is comprised of mostly dark-colored Australian species (e.g., P. interrupta ), which often have a wide or moderately wide (but sometimes also rather narrow) forewing, a relatively short to moderately long female antenna, and a relatively short and weak axillar seta. The fourth species group unites the mostly undescribed, peculiar species similar to P. sappho , which often have colorful bodies and various dark markings on the forewing (Figs 79, 80); we have seen species belonging to this group from Queensland ( Australia), Papua New Guinea, Fiji, and also the high elevation, native forests of French Polynesia.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Palaeoneura Waterhouse 1915
| TRIAPITSYN, SERGUEI V. & BEREZOVSKIY, VLADIMIR V. 2007 |
Acanthomymar
| Huber, J. T. 2003: 80 |
| Huber, J. T. 2003: 80 |
| Hayat, M. & Anis, S. B. 1999: 307 |
| Subba Rao, B. R. 1970: 668 |
Chaetomymar
| Huber, J. T. 2003: 78 |
| Annecke, D. P. & Doutt, R. L. 1961: 34 |
| Ogloblin, A. A. 1946: 277 |
Palaeoneura
| Annecke, D. P. & Doutt, R. L. 1961: 30 |
| Soyka, W. 1956: 107 |
| Gahan, A. B. & Fagan, M. M. 1923: 103 |
| Waterhouse, C. O. 1915: 538 |