
Careproctus scottae Chapman & DeLacy 1934
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4018.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:BDEF6499-7A88-496E-AB24-A82977F0D1A5 |
|
DOI |
https://doi.org/10.5281/zenodo.5618217 |
|
persistent identifier |
https://treatment.plazi.org/id/D33887A3-FFB6-FF98-FF3E-FAD5FBD0FBC9 |
|
treatment provided by |
Plazi |
|
scientific name |
Careproctus scottae Chapman & DeLacy 1934 |
| status |
|
Careproctus scottae Chapman & DeLacy 1934 View in CoL
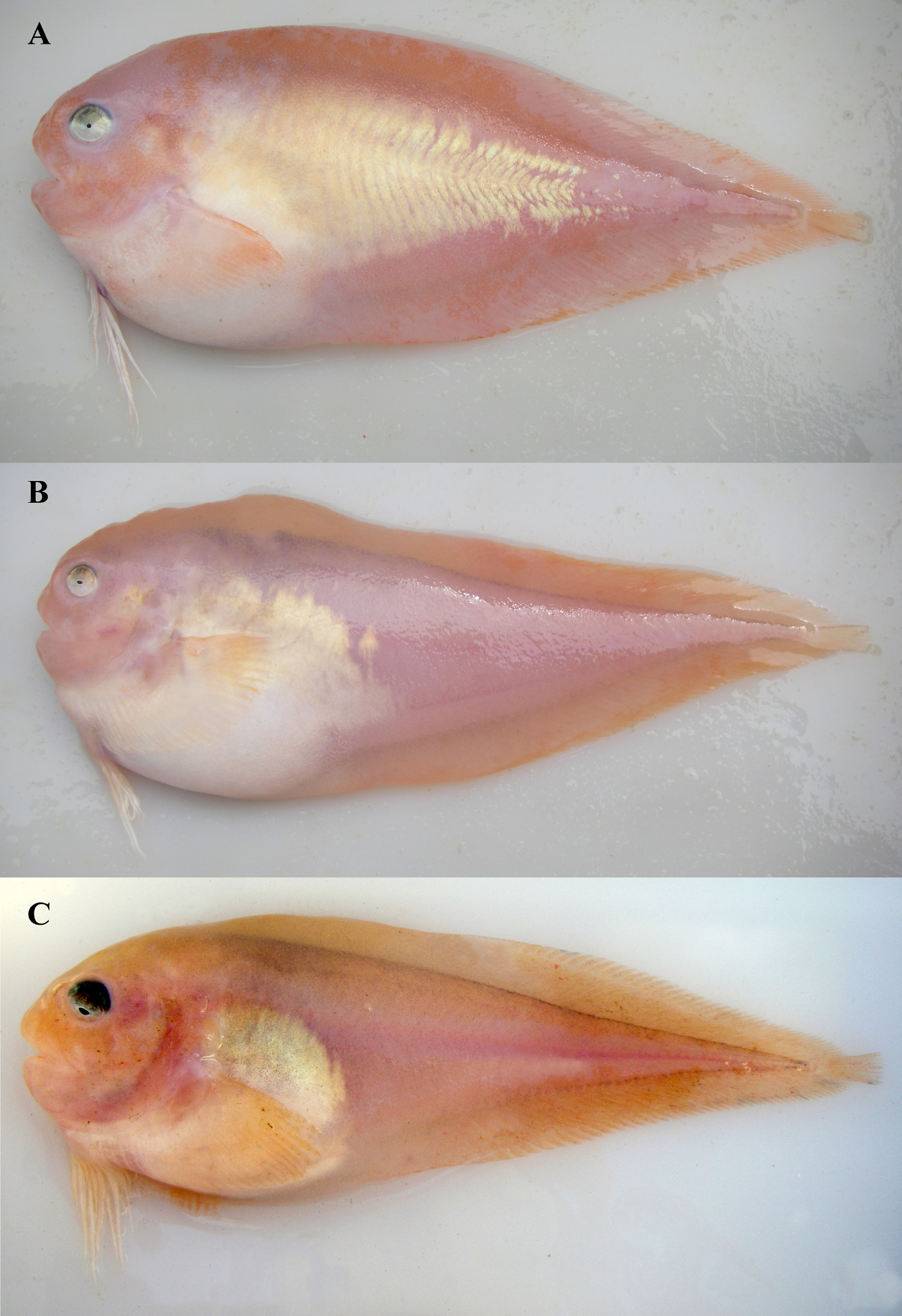
English Common Name: Peachskin Snailfish Figures 3 View FIGURE 3 A, 5C, 6; Tables 1–3
Careproctus scottae Chapman & DeLacy 1934:2 View in CoL . Type locality: southeast Alaska, off Petersburg.— Taranetz 1937:137 (list, Alaska).— Quast & Hall 1972:29 (Alaska).— Allen & Smith 1988:67 (Alaska, “pink snailfish”, in part).—Robins et al. 1991:44 (list).— Sheiko & Fedorov 2000:32 (listed as questionable synonym of C. rastrinus View in CoL ).— Mecklenburg et al. 2002:612 (Alaska, in key, listed as probable synonym of C. rastrinus View in CoL ).— Chernova et al. 2004:16 (checklist).— Nelson et al. 2004:124 (list), Love et al. 2005:103 (checklist).— Kai et al. 2011a:144 (genetics, morphology, phylogenetics, as “BER1” and “BER2”).— Kai et al. 2011b:368 (fig. 1a, phylogenetics, as “BER1” and “BER2”).—Page et al. 2013:127 (list).
Holotype. USNM 104497 [ex UW 2742], 187.7 mm, southeast Alaska, off Petersburg, Dorothy R. Scott.
Paratypes. UW 2741, 5 (123.0–180.0 mm), Alaska, Frederick Sound, Thomas Bay, 57ºN, 132.783ºW, 183 m depth, 1920; UMMZ 102220, 1 (143 mm), off Petersburg, 31 December 1932; USNM 93787, n = 1, radiograph only, off Petersburg, January 1933; UW 2743, n = 2, and USNM 104681 [ex UW 2743], n = 3, off Petersburg, 20 June 1932 [not seen]. All collected by Dorothy R. Scott.
Additional material examined. A total of 344 specimens, not including the types above, 44.6–253.2 mm SL. See “Non-type material examined below.”
Diagnosis. Careproctus scottae is distinguished from all other species of Careproctus by the combination of cyt b and 16S rRNA sequence data (“BER1” and “BER2” of Kai et al. 2011a; Table 1), an anteriorly robust body covered by cactus-like prickles, the presence of the postorbital pore, a moderate-sized pelvic disc, a lower pectoralfin lobe longer than upper lobe, a pale peritoneum and stomach, and counts of vertebrae and median fin rays. It is most similar to C. rastrinus , from which it can be distinguished by its lower counts of caudal vertebrae (48–52 in C. scottae vs. 51–56 in C. rastrinus ) and anal-fin rays (47–53 vs. 51–56), pale peritoneum and stomach (gray in C. rastrinus ), a larger pelvic disc in individuals larger than 100 mm (13.9–28.8 vs. 9.9–18.8 % HL), and a longer nasal tube (3.2–6.4 vs. 0.9–4.7 % HL), and to C. phasma , from which it can be distinguished by the postorbital pore (present in C. scottae vs. absent in C. phasma ), smaller pelvic disc (13.9–28.8 vs. 20.1–35.9 % HL), higher counts of dorsal-fin rays (54–61 vs. 50–56), higher counts of anal-fin rays (47–53 vs. 43–49), and higher counts of vertebrae (59–64 vs. 55–60). It is further distinguished from C. spectrum by its higher counts of dorsal-fin rays (54–61 vs. 52–54 in C. spectrum ) and smaller orbit (16.4–32.1 vs. 33.9–34.4 % HL).
Description. Body heavy and deep anteriorly, tapering strongly posteriorly, strongly compressed; depth at pectoral-fin base 81.0–154.8 (104.7) % HL. Head large, 21.7–32.0 (26.0) % SL, robust, dorsal profile rounded from nape to snout. Snout blunt, slightly projecting anterior to lower jaw. Mouth terminal, small, horizontal; upper jaw 35.9–57.3 (39.1) % HL, maxilla extending to anterior part of orbit or mid-orbit, oral cleft extending to anterior rim of orbit; mandible 45.1–63.0 % HL. Premaxillary tooth plates matching mandibular tooth plates. Premaxillary and mandibular teeth simple with weak shoulders in 20–38 oblique rows of 6–11 teeth forming narrow bands. Diastema absent at symphysis of upper and lower jaws. Orbit 16.4–32.1 (20.3) % HL, dorsal margin well below dorsal contour of head, suborbital depth to upper jaw 8.4–18.9 (18.5) % HL, to lower jaw 27.5–36.0 (31.8) % HL; pupil round. Interorbital space broad, fleshy distance 20.3–62.9 % HL, bony distance 18.2–34.5 (29.2) % HL, strongly convex. Snout much longer than orbit, 102.8–244.1 (188.7) % OL, 28.1–47.5 (38.2) % HL. Nostril single, with well-developed tube at level with lower rim of orbit; nostril tube length 5.8–19.7 % OL.
Pores of cephalic lateralis system of moderate size, pore pattern 2-6-7-2, chin pores paired. Interorbital pore absent.
Gill opening small, 18.3–44.2 (30.1) % HL, upper margin at or just above level of dorsal rim of orbit, extending ventrally to above pectoral-fin or to pectoral-fin rays 1–8 (ray 3). Opercular flap rounded to slightly angular (rounded). Gill rakers 8–12 (Tables 2–3), short, blunt.
Dorsal-fin rays 54–61 (56; Tables 2–3), anterior dorsal lobe absent, anterior rays buried in tissues, tips of more posterior rays not exserted. Anteriormost dorsal-fin pterygiophore inserted between neural spines 2 and 3 or 3 and 4, rayless or bearing a single small ray (3 and 4, rayless). Predorsal length 22.8–34.8 (28.7) % SL. Anal-fin rays 47–53 (50; Tables 2–3), zero to three (two) anal-fin pterygiophores anterior to first haemal spine, each bearing a single ray, tips of all rays not exserted. Anal-fin origin below vertebrae 13–14 (caudal vertebrae 2–3), preanal length 30.5–49.5 (38.5) % SL.
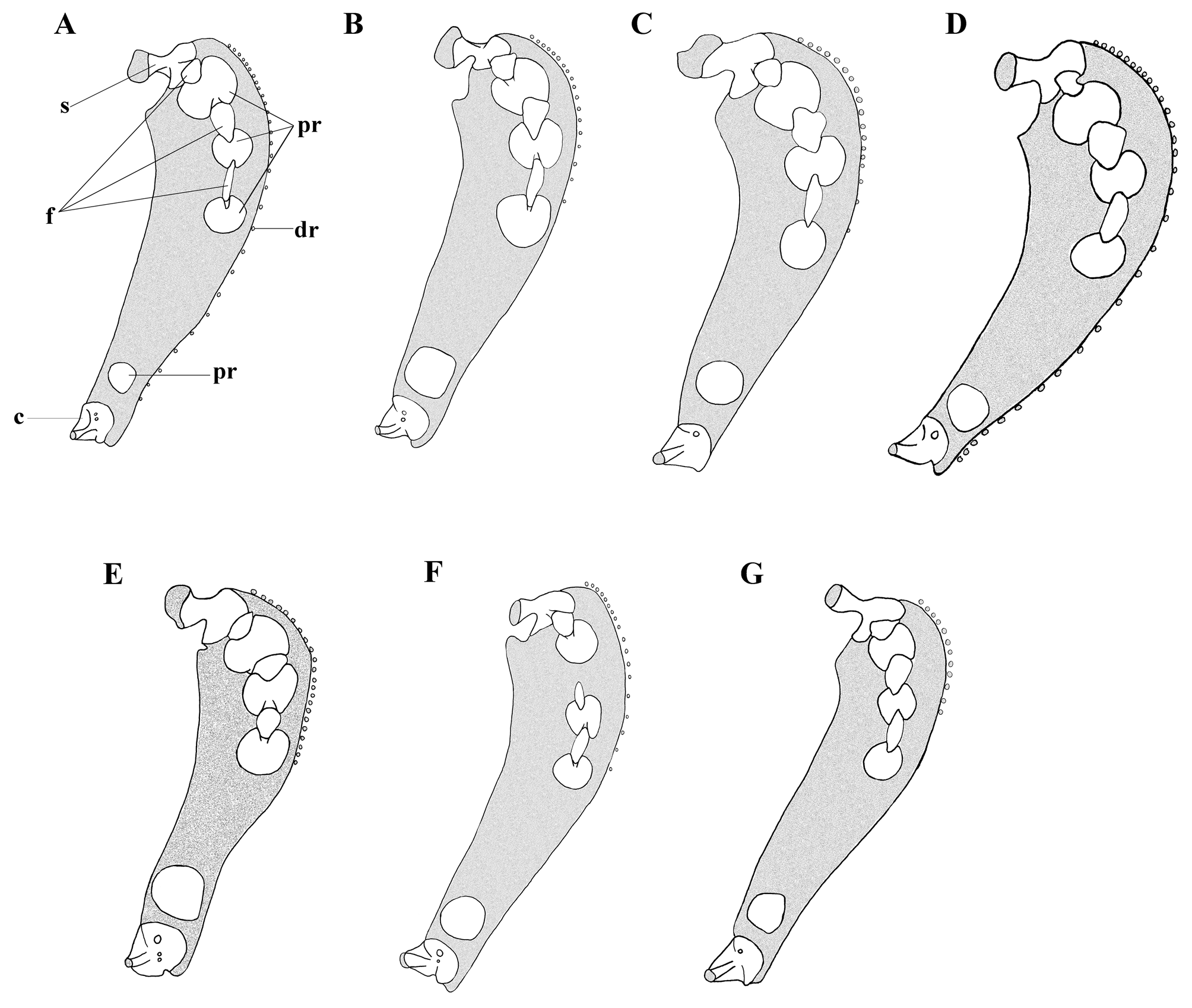
Pectoral fin deeply notched, with 31–39 (32) rays (Tables 2–3). Upper lobe 46.9–75.6 (61.2) % HL, with 21– 32 (24) rays extending well beyond anus to or near anal-fin origin, shorter than lower lobe, dorsalmost rays lengthening to rays 8–10, more ventral rays gradually shortening to shortest ray of notch. Lower lobe elongate, 54.2–1.33 (98.3) % HL, with 7–12 (8) rays, extending well beyond anus to or near anal-fin origin; dorsal rays gradually shortening to ventralmost ray near pectoral symphysis. Tips of rays 5–20% free of membrane, elongate rays of lower lobe mostly free of membrane. Notch strong, rays in notch slightly more widely spaced than rays of lobes, more widely spaced ventrally. Uppermost pectoral-fin ray level with region between ventral rim of orbit and oral cleft. Insertion of lowermost pectoral-fin ray below mid-orbit. Proximal pectoral radials four (3+1), robust: radials 1–2 notched and hour-glass shaped, radial 3 round, with slight dorsal notch, radial 4 round ( Fig. 5 View FIGURE 5 C). Interradial fenestrae three, extending between scapula and proximal radials 1–3: fenestrae between the scapula and radials 1 and 2 elliptical, fenestra between radials 2 and 3 narrow and elongate. Scapula broadly T-shaped with robust distally broadened helve; coracoid with broad triangular head and broad helve, angled slightly anteriorly. Distal radials present at base of rays 2–19, ventralmost at level of proximal radial 3, dorsalmost ray and more ventral rays articulating directly with pectoral cartilage.
Pelvic disc small, length 13.9–26.6 (16.4) % HL, round, about as long as wide, width 11.2–26.1 (16.7) % HL, anterior lobe weakly developed, slightly to strongly cupped, distance from tip of snout to pelvic disc 9.9–15.8 % SL. Anus below posterior rim of orbit, close behind pelvic disc; distance from snout to anus 13.1–26.1 (18.4) % SL, 50.2–96.5 (70.8) % HL.
Principal caudal-fin rays 9–10, dorsal procurrent rays 1–3, ventral procurrent rays 0–2 (1–3 + 4–5/5 + 0–2) (? + 4/5 +?). Caudal fin 32.3–48.7 (41.1) % HL. Membrane of posterior dorsal-fin rays attached to caudal fin at shorter distance than anal-fin rays: dorsal-fin rays attached to caudal fin 26.9–64.0 (53.7) % CL; anal-fin rays, 43.4–79.3 (48.0) % CL. Depth at base of caudal fin 13.4–29.2 (22.6) % CL.
Skin relatively thick, thick gelatinous layer beneath skin, cactus-like prickles uniformly covering body, in most dense region about 13 prickles in orbit length. Pyloric caeca 15–22, length about 32–64 (53.2) % HL, center-left side of visceral cavity.
Vertebrae 59–64 (62), precaudal 10–13 (11), caudal 48–52 (51; Tables 2–3). Pleural ribs 2 or 3 (2), anteriormost small when 3, others long and slender, present on vertebrae 9–10 or 8–10 (9–10).
Coloration. Body and fins orangish pink and white in life ( Fig. 3 View FIGURE 3 A); head, dorsum from nape to caudal fin, and ventrum at anal-fin origin to caudal fin orangish pink; isthmus, base of pectoral fin, and body posterior of gill slit to anal-fin origin white; area above belly silvery white (crystalline guanine) with orangish-pink highlights becoming obsolete posteriorly at about a quarter to half the anal-fin length; base and lower lobe of pectoral fin white; dorsal margin and distal portion of fin orangish pink; eye dark dorsally, silvery greenish gray ventrally. Body and fins pale in preservation; base of fins beneath skin with pigment, showing faint line between fins and body. Peritoneum pale to lightly speckled; orobranchial cavity pale; stomach pale to gray, intestines pale to mottled, pyloric caeca pale to mottled, and urogenital papilla pale.
Life history. The largest specimen examined was 253.8 mm (UW 117348), a ripe female with yolked eggs, and the largest specimen observed was about 390 mm from off Amchitka Island in the Aleutian Islands (AFSC 2000 survey photograph, JWO, unpublished data). The smallest ripe female with yolked eggs was 186.8 mm. Ripe yolked eggs were about 2.6 mm in diameter. The smallest ripe male was 123.9 mm; the largest, 237 mm.
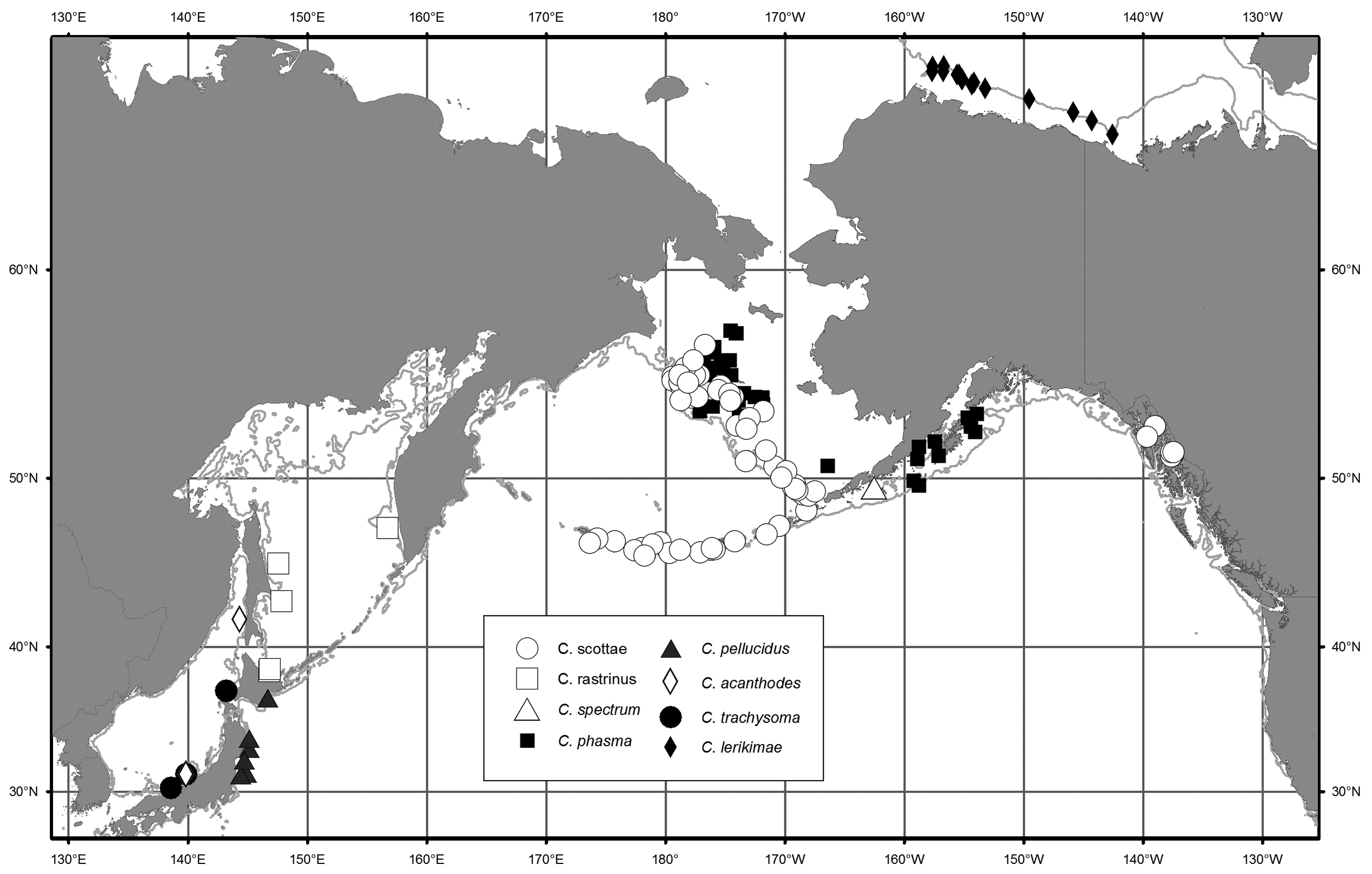
Distribution. Careproctus scottae has been collected throughout the deeper waters of the eastern Bering Sea and the Aleutian Islands ( Fig. 6 View FIGURE 6 ). Collection depths range from 71 to 390 m. Previously recognized as C. rastrinus outside of Southeast Alaska, it is one of the most commonly identified species of Careproctus on the Bering Sea slope (e.g., Hoff 2013) and in the Aleutian Islands (e.g., Von Szalay et al. 2011). Records of C. rastrinus from the western Bering Sea (e.g., Orlov 2005) may be either C. scottae or C. rastrinus and require verification.
Etymology. The specific epithet is a patronym honoring the collector of all the type material, Dorothy R. Scott.
Remarks. Kai et al. (2011a) identified two closely related clades (BER1 and BER2) within C. scottae found sympatrically in the Bering Sea. These clades may be deserving of species-level recognition based on the similar level of sequence divergence found among other species of this complex. However, no difference in morphology was detected between specimens genetically assigned to these clades. Additional material from clade BER2 or nuclear DNA sequence data (e.g., Kai et al. 2011b) may be required to clarify its status.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Careproctus scottae Chapman & DeLacy 1934
| Orr, James Wilder, Kai, Yoshiaki & Nakabo, Tetsuji 2015 |
Careproctus scottae
| Kai 2011: 144 |
| Kai 2011: 368 |
| Chernova 2004: 16 |
| Nelson 2004: 124 |
| Mecklenburg 2002: 612 |
| Sheiko 2000: 32 |
| Allen 1988: 67 |
| Quast 1972: 29 |
| Taranetz 1937: 137 |
| Chapman 1934: 2 |