
Petrolestes hendersoni Cockerell, 1927
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4028.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:1F5BEBE1-4BAE-4EB4-8BDE-ACA0FF2D1866 |
|
DOI |
https://doi.org/10.5281/zenodo.6105804 |
|
persistent identifier |
https://treatment.plazi.org/id/CF6C9266-AA60-5252-17AB-62D9FD41FCFB |
|
treatment provided by |
Plazi |
|
scientific name |
Petrolestes hendersoni Cockerell, 1927 |
| status |
|
Petrolestes hendersoni Cockerell, 1927
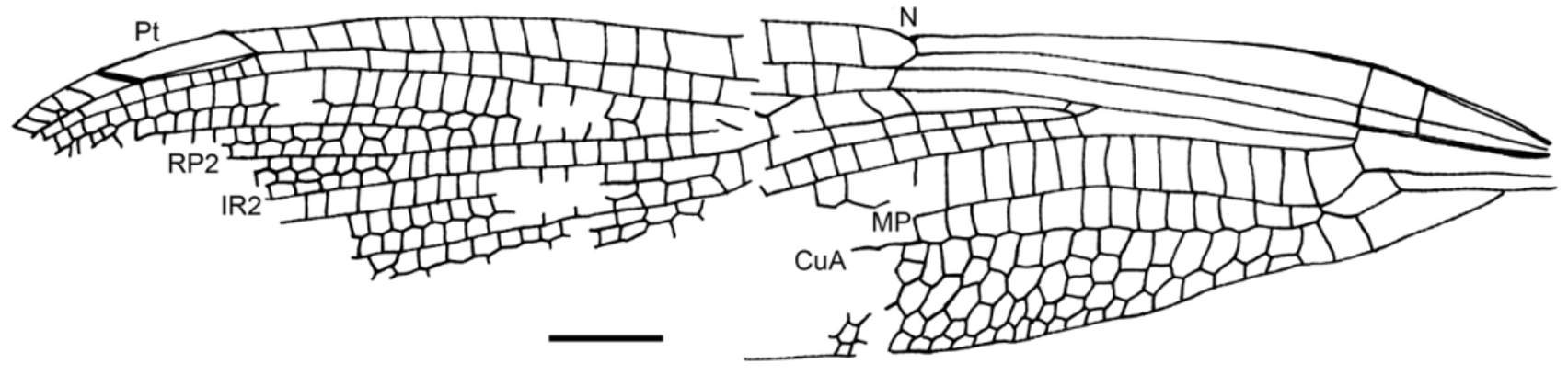
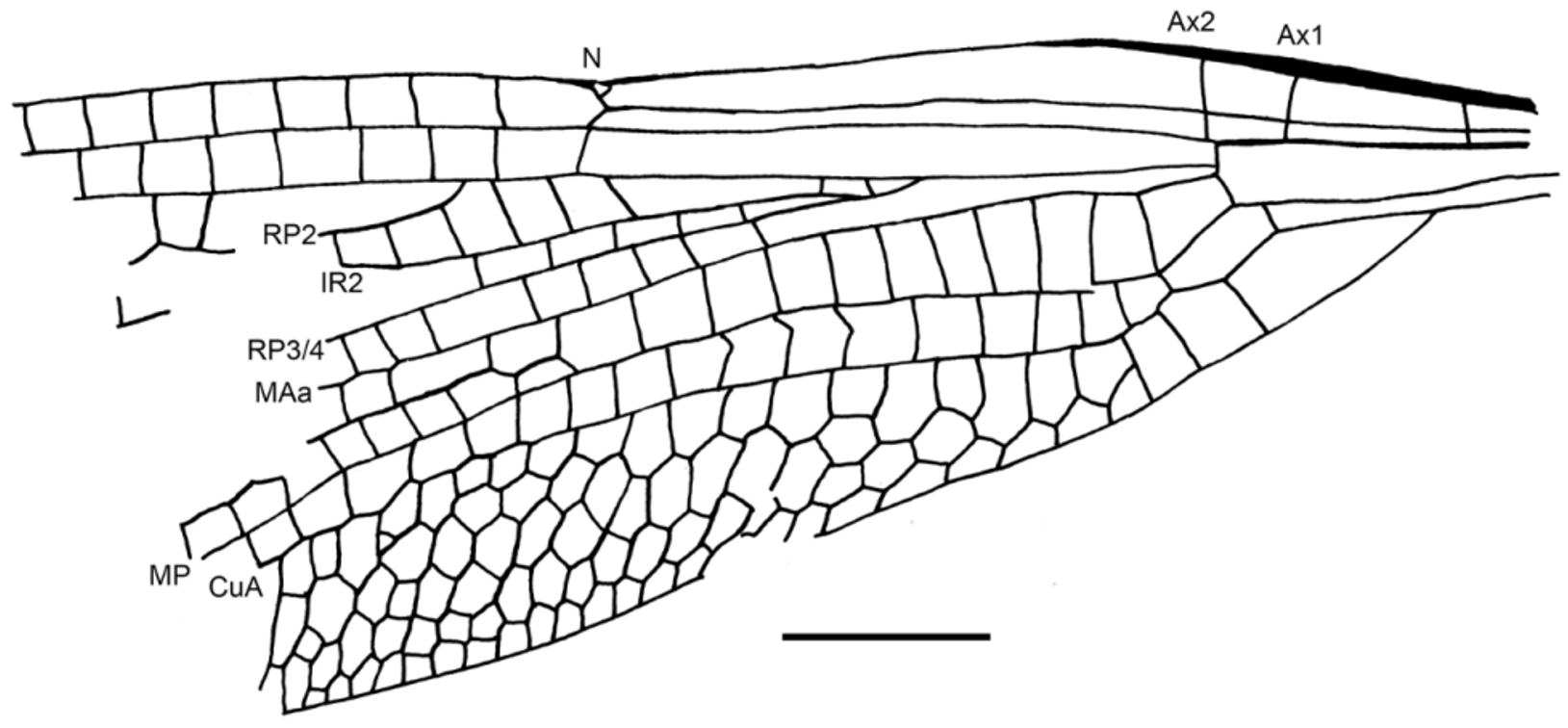
( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Material. holotype specimen UCM 15619, University of Colorado, Boulder, Geology Section, Museum Annex, USA; new specimen USNM without number, Smithsonian Institution, National Museum of Natural History, USA. Age and outcrop. Eocene, Green River Formation; the type specimen UCM 15619 comes from the Ute Trail, Roan Plateau, Garfield County; the new specimen USNM without number comes from Wighton Garfield County; both in Colorado, USA.
Emended diagnosis. Wing characters only. Intercalaries between MA and MP present just distal of nodus level; 17 postnodals; wing rather large, 8.0 mm wide at nodus level.
Description of new specimen. Body with head and five basal segment of abdomen attached to thorax, together with the four wings; only the left forewing is nearly complete, basal half of left hind wing and basal fourth of right wings preserved; no trace of secondary genital structures on abdominal segment 2 (female specimen).
Head rather broad, 3.6 mm long, 5.2 mm wide, eyes 3.0 mm wide, widely separated, 2.4 mm apart; ocelli not discernable; thorax 6.0 mm high, moderately prognathous, thoracic skewness (sensu Needham and Anthony, 1903) moderate, comparable to that of a modern Aeshnidae , angle x 35 °, angle z ca. 30°; interpleural suture (sensu Asahina 1957) incomplete but formed at dorsal 1/2; abdomen 2.0 mm wide. Legs robust, with tibia spines strong, slightly longer than distance between two spines.
Wings hyaline, forewing ca. 40.0 mm long; max. width 8.5 mm, wing 8.0 mm wide at nodus level; a very short petiole, 1.5 mm long; anal area with one row of cells between AA and AP; subdiscoidal cell broad elongate, 1.8 mm long, 0.8 mm wide; distance from base to arculus 5.0 mm, from arculus to nodus 11.2 mm, from nodus to pterostigma 16.5 mm; nodus in a basal position; a significant increase of spine density at the apical costal margin; pterostigma long and broad, 4.0 mm long, 0.8 mm wide, covering six cells; pterostigmal brace perpendicular to RA and RP 1; distance from wing base to Ax1 3.8 mm, from Ax1 to Ax2 1.5 mm, Ax2 nearly aligned with arculus; no supplementary antenodal crossvein; discoidal cell unicellular, 1.4 mm long, 1.4 mm wide, distinctly broadened in its distal part, basal side 0.8 mm, anterior side 1.2 mm, posterior side 1.4 mm, distal side MAb 1.4 mm; ScP not crossing through nodus as in Stenolestes Scudder, 1895; nodal crossvein very oblique; subnodus slightly oblique with corresponding crossvein below (between RP 1/2 and IR2) of inverted obliquity; ca. 17 postnodal crossveins preserved, not aligned with the 16 postsubnodal crossveins; bases of RP 3/4 and IR2 between arculus and nodus, closer to nodus than to arculus, base of RP 3/4 6.5 mm from arculus and 4.7 mm from nodus; base of RP 2 two cells, 2.2 mm distal of subnodus; base of IR1 four cells and 4.0 mm distally; oblique crossvein ‘O’ absent; cubito-anal area with five rows of cells between CuA and posterior wing margin; CuA reaching posterior wing margin well distal of nodus level; CuA weakly curved; postdiscoidal area with one row of cells up to nodus level and two rows more distally; area between RP 3/4 and MA with at least two secondary longitudinal veins in between; three rows of cells between RP 3/4 and IR2 and one row between RP 1 and IR1; three rows of cells between IR1 and RP 2 opposite pterostigma, but only one basally.
Preserved part of hind wing nearly identical to forewing, even the shape of the discoidal cell nearly the same. Only the position of the nodus is slightly different, as distance from arculus to nodus is 8.7 mm.
Discussion. The comparison of the preserved parts (hind wing basal two-thirds) of the venation of the type of Petrolestes hendersoni with that of this new fossil shows that they are nearly identical in every point (petiole length, nodus, subnodus, discoidal cell, antenodal and cubito-anal areas, relative positions of bases of branches of RP, number of rows of cells between main veins in basal part of wings). Also the dimensions of the wing structures are nearly identical in the two specimens. As both specimens come from the Green River Formation in Colorado and even from outcrops in the same county, we consider that they belong to the same species. The new specimen allows more precision in several points concerning this taxon, viz. it has: no oblique vein ‘O’, a long pterostigma covering numerous cells, a long zigzagged IR1, a short petiole, and the discoidal cells of the fore and hind wings are nearly identical. These characters confirm the position of Petrolestes close to Congqingia Zhang, 1992 in the Dysagrioninae, rather than close to the Sieblosiidae . In particular Petrolestes has not the subnodal veinlet vertical or anteriorly oriented and its nodus is not traversed by ScP, unlike in the Sieblosiidae ; it also has no oblique vein ‘O’. Petrolestes has also a significant increase of spine density at the apical costal margin, a character proposed by Bechly (1996) as a potential synapomorphy of the Zygoptera, absent in the Sieblosiidae ( Fleck et al. 2004) . Petrolestes and Congqingia share with Dysagrion a rather unique shape of the discoidal cell, potential synapomorphy even if it is also present in the Sieblosiidae . The presence of a ‘basal costal margin between wing base and nodus distinctly convex curved’ (a character absent in the Sieblosiidae , see Nel et al. 2005), plus the ‘antesubnodal space without any crossveins’ (a character present in many Zygoptera and in the Sieblosiidae ) would be the unique putative synapomorphies of the Dysagrioninae with the modern Thaumatoneuridae , after Bechly (2014). If Dysagrion shares with Thaumatoneura similar relative positions of the nodus and bases of the branches of RP, it is not the case for Petrolestes and Congqingia, which have the bases of RP 3/4 and IR2 basally recessed midway between the nodus and the arculus. Also the discoidal cell of Thaumatoneura is elongate and much narrower than in all the taxa currently included in the Dysagrioninae. Also Thaumatoneura has long spines on its legs, unlike Petrolestes, and its thoracic skewness is much more pronounced. Thus the attribution of this group of damselflies as sister group of the modern Thaumatoneuridae remains debatable.
Several characters suggest that Petrolestes (and consequently Congqingia and maybe Dysagrion) could not be a Zygoptera: the head appears not as transverse as in a Zygoptera (but this could be due to the compression of the fossil that can strongly deform the head shape; similar situations occur for fossil Lestes in compression on rocks from the Oligocene of France, so that the head seems to be less transverse than it was certainly in the living animal); the thorax is not very skewed, similarly to what happen in Epiproctophora (but this could be due to the robustness of this animal); the legs have spines shorter and stronger than in Zygoptera; this character is rather present in Epiproctophora. Also the position of the arculus near Ax2 instead of being midway between Ax2 and Ax1 or closer to Ax1 is a plesiomorphic character that would exclude a position within the Epiproctophora for Petrolestes. The presence of an incomplete interpleural suture but rather long and formed at dorsal ½ is a plesiomorphic character present in Zygoptera (the interpleural suture is complete in Thaumatoneura ) but not in Epiproctophora.
It remains that the thoracic skewness and relatively short leg spines of Petrolestes suggest affinities with the Epiproctophora rather than with Zygoptera. Petrolestes, and consequently Congqingia and Dysagrion, could be taxa in the stem lineage of the Epiproctophora rather than Zygoptera. It would be more accurate to have information on the genital appendages of these damselflies before an attribution close to the Epiproctophora.
Petrolestes differs from Congqingia in the longer pterostigma, covering six cells, instead of two, in the broader area between IR1 and RP 2 with longer intercalary veins between them, and in the broader cubito-anal area with five-six rows of cells instead of two (Zhang 1992).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |