
Chonocephalus Wandolleck, 1898
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1772.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5123987 |
|
persistent identifier |
https://treatment.plazi.org/id/C36887E2-8727-FF8B-AD94-FD21FBC7FF74 |
|
treatment provided by |
Felipe |
|
scientific name |
Chonocephalus Wandolleck, 1898 |
| status |
|
Genus Chonocephalus Wandolleck, 1898 View in CoL View at ENA
Type species. Chonocephalus dorsalis Wandolleck, 1898: 428 View in CoL .
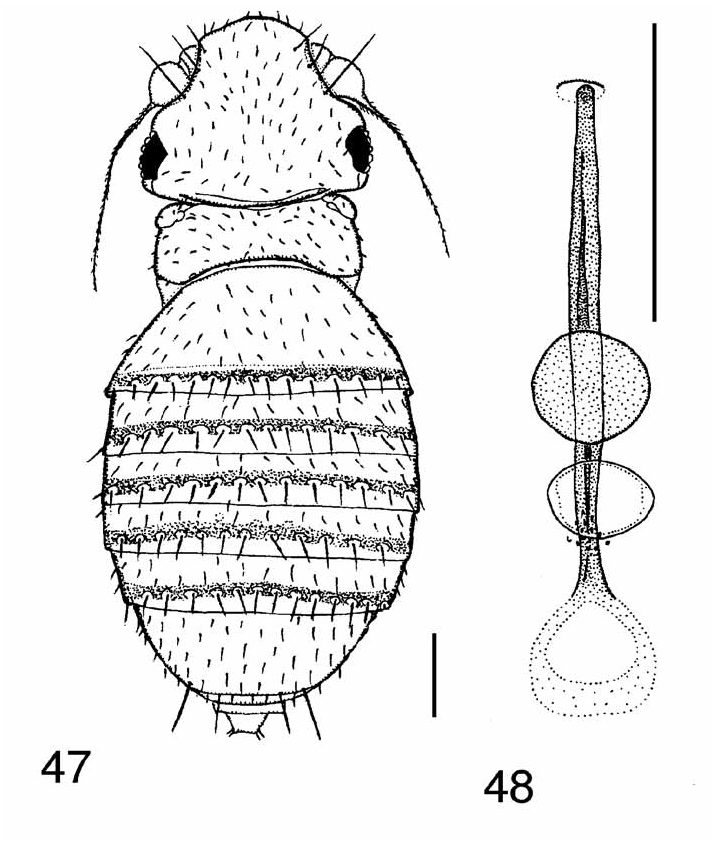
The genus is readily identified in the most recent key to world genera ( Disney, 1994), with the modification that the genus Epichonocephalus Schmitz View in CoL has been synonymised with Chonocephalus ( Disney, 2002) View in CoL . The same paper provides a detailed diagnosis of the genus in both sexes. The wings of the males are distinctive ( Figs 18 View FIGURES 15–18 and 64 View FIGURES 64–65 ) as is the general appearance of the females ( Figs 47 View FIGURES 47–48 and 66 View FIGURES 66–68 ).
The full details of the references for synonyms, etc., covered by Disney (2002) are not repeated in this paper.
Males. The recognition of species depends primarily on the details of the hypopygium, which is at first sight very complex. However, if one ignores the, often elaborate, penis complex (X), the details of the epandrium (E) and hypandrium (H) that are referred to in the key below are indicated in Fig. 1 View FIGURE 1 . This represents a generalised, stylized, hypopygium. While the anal tube (A) is seldom utilised in the recognition of species it is a useful point of reference. Any one of the labelled structures may be modified and may sometimes be very elaborate in form. In the inferred groundplan state the rear margin of the hypandrium bears four processes. These are the gonopods (G) and a pair of median lobes (L). In the plesiomorphic state the gonopods articulate with the dorsal face of the hypandrium, but more frequently they arise from it. One (usually the left) or both may be reduced or absent. The median lobes (L) may form a single bilobed structure or be reduced to a single lobe (which may be greatly elongated) or be lost altogether. Ventrally each side of the epandrium (E) typically bears an anterior process (AP) and a posterior process (PP). In some species the anterior processes (AP) may appear to be appendages (attaching to the inner face of the epandrium) rather than simple projections of the epandrium. A presumed true left surstylus or clasper (C) is frequently present (e.g. Figs 17 View FIGURES 15–18 and 24 View FIGURES 24– 26 ), variably fused to the epandrium, and is sometimes elaborate. Occasionally it is fused to the bridge (B), which is sometimes present and probably represents sternite 10. The most anterior (lower) bristle of the left side of the epandrium is sometimes modified into a spine (S), which may be further modified.
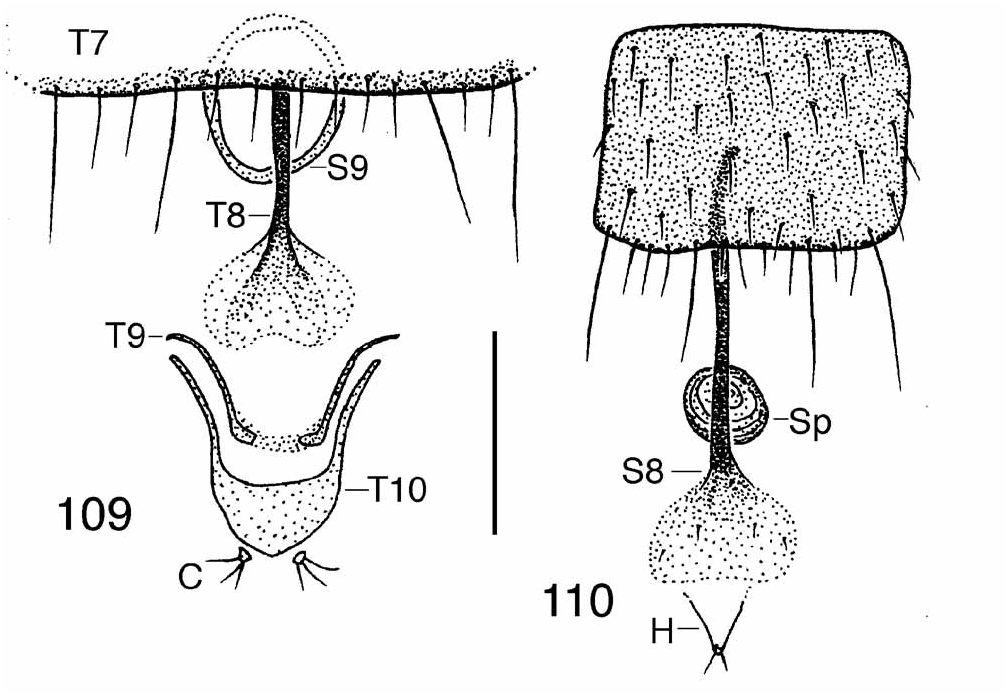
Females. The critical details of the terminal segments of the abdomen are indicated in Figs 109–110 View FIGURES 109– 110 , which depicts a basic (close to the groundplan) state of a female Chonocephalus ( Disney, 1986) View in CoL . In most species the T9 is lost apart from its anterolateral apodemes, which are fused to those of T10 so that the latter appear to be two-branched.
Review of species. The descriptions of the new species are brief as the significant features of the males are largely restricted to the details of the hypopygia. Likewise in the females there are relatively few features that distinguish the species apart from the details of the abdominal terminalia. The formal diagnoses, therefore, are the features used to distinguish the species in the keys above.
The brief notes on the natural histories of the species, when known, are taken from Disney (2002) except where indicated otherwise. Typical larvae and pupae are shown in Figs 70–74 View FIGURES 70–71 View FIGURES 72–74 .







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.