
Limnonectes sisikdagu, Mcleod, David S., Horner, Stephani J., Husted, Colin, Barley, Anthony & Iskandar, Djoko, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.201604 |
|
DOI |
https://doi.org/10.5281/zenodo.5618573 |
|
persistent identifier |
https://treatment.plazi.org/id/BD152C78-FFA6-006C-65D5-F8EA17A4F9A4 |
|
treatment provided by |
Plazi |
|
scientific name |
Limnonectes sisikdagu |
| status |
sp. nov. |
Limnonectes sisikdagu View in CoL sp. nov.
Holotype. Adult male ( FMNH 266636; Fig. 2 View FIGURE 2 ), from Batang Andaleh, Lubek Selasih near Solok at 01˚0'37"S 100˚37'55"E, 1204 m to 01˚0'33"S 100˚38'20"E, 1252 m, collected on 0 6 September 2001 by Djoko Iskandar.
Paratypes. Paratypes collected by Djoko Iskandar and deposited at the Field Museum of Natural History. Four adult males ( FMNH 266619, 266630–31, 266637), four adult females ( FMNH 266608, 266610, 266615, 266618), two juveniles ( FMNH 266619, 266621). FMNH 266608 and FMNH 266610 (females), collected from Batang Harau, near Harau Valley, Payakumbuh at 00˚4'06"S 100˚39'17"E, 536 m to 00˚6'42"S 100˚39'51"E, 566 m on 07–08 August 2001. FMNH 266615, 266618 (females), 266616 (male), 266619 and 266621 (juveniles) were all collected from Sarasah Bunta, near Harau Valley, Payakumbuh at 00˚5'37"S 100˚40'15"E, 523 m to 00˚6'42"S 100˚40'28"E, 530 m on 08–17 August 2001. FMNH 266630 and 266631 (males) were collected at Batang Tarusan, Lubuk Selasih, near Solok at 01˚0'44"S 100˚37'55"E, 1170 m to 01˚54'42"S 100˚28'19"E, 1166 m on 02–10 September 2001. FMNH 266637 (male) was collected with holotype.
Referred specimens. Collected by David Bickford and deposited at the Raffles Museum of Biodiversity Research ( RMBR). RMBR 393 ( HM067244 View Materials ) from Batu Layng 03°27'49"S 102°18'83"E, 410m on 19 July 2007; RMBR 515 ( HM067245 View Materials ) from Batu Layng 03°27'57"S 102°19'10"E, 460m on 21 July 2007. Represented in this study and McLeod (2010) by sequence data derived from tissue samples. Whole, preserved voucher specimens unavailable for examination.
Diagnosis. Limnonectes sisikdagu is considered a member of Limnonectes on the basis of molecular evidence and the following morphological diagnostic characters: presence of fanglike odontoid processes on the lower jaw (a sexually dimorphic feature; larger in males than females); male-biased size dimorphism (generally characterized by the hypertrophy of the head in males) ( Emerson et al., 2000). Additionally, these specimens are part of the L. kuhlii Complex on the basis of molecular data, and possession of an indistinct (or hidden) tympanum and fully webbed toes, both of which are characters traditionally used to diagnose L. kuhlii sensu lato from its congeners ( Boulenger, 1920; Duméril & Bibron, 1841; Inger, 1966; Taylor, 1962; Tschudi, 1838).
Limnonectes sisikdagu can be distinguished from true L. kuhlii (Java) , and from other members of the kuhlii Complex, by the following combination of characters: (1) males with spinule-covered mental patch ( Fig. 3 View FIGURE 3 ); (2) adult male SVL 41.84–49.43 mm (mean = 45.36; SD ± 3.22; n = 5), adult female SVL 44.44–56.63 mm (mean = 49.10; SD ± 5.50; n = 4); (3) males with nuptial pads on first finger (Finger II; Fig. 4 View FIGURE 4 ); (4) males with moderately enlarged heads (HL 43.84–47.97% of SVL; 39.43–42.76% in females); (5) males and females with heads that are longer than wide (HL 107–114% HW in males; 108–112% in females); (6) prominent post-orbital fold; (7) discontinuous dorsolateral fold; (8) throat lightly pigmented (mottled), moderate pigmentation on borders of thigh and leg, venter and ventral thigh immaculate; (9) moderately dense small, low glandular warts tipped with transclucent spinules on flanks of body, around vent and on dorsum of thigh, shank, and foot; (10) dorsal skin and throat feebly crenulate; (11) relative finger length (longest to shortest) when adpressed: IV–V–II–III.
Description of holotype. Adult male ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). Habitus robust with moderately enlarged head (HL 48% SVL); head longer than wide (HL 109% HW). Rostrum pointed in dorsal view, projecting beyond lower jaw, obtuse (sloping) in profile; nostril dorsolaterally oriented, closer to tip of snout than to eye; internarial distance greater than interorbital distance; canthus rounded; lores concave; upper lip distinctly swollen and flared, not reaching post-rictal tubercle; eye diameter 23% head length; upper eyelid width greater than interorbital distance. Supratympanic fold moderate, extending from eye to angle of jaw (insertion of arm); tympanic annulus not visible through skin; post-orbital skin fold intersects supratympanic fold just posterior to eye. Vomerine teeth on oblique ridges, separated from each other by much less than width of one ridge. Choanae oval, perpendicular to longitudinal axis of body. Odontoid processes robust with rounded tips, length nearly twice depth of mandible at base of processes. Symphysial knob at mandibular symphysis. Tongue oval, deeply notched posteriorly. Mental patch on anterior portion of chin covered with small translucent spinules, larger than, but resembling, those on nuptial pads.
Finger tips rounded, not expanded into discs, with rounded distal pad; decreasing lengths of fingers: IV–V–II– III; no webbing between fingers; distinct, movable fringe of skin on pre-and postaxial sides of Fingers III and IV; digits indicated by Roman numeral (tubercle count in parentheses): IV (2), III (2) II (1), I (1); proximal subarticular tubercles prominent, round, elevated on Fingers III and IV; distal subarticular tubercles low, flat and indistinct; thenar metacarpal tubercle large, oval, not elevated; inner metacarpal tubercle oval, smaller than thenar tubercle, not contacting outer or thenar tubercles; outer metacarpal tubercle smaller than inner tubercle, oval, elevated; prominent nuptial pad composed of minute spines on medial surface of Finger II from distal subarticular tubercle along penultimate distal phalanx and in a small patch on the thenar metacarpal tubercle. Tips of toes rounded, not expanded into discs, toe pads elevated; decreasing lengths of toes: IV–III–V–II–I; toes webbed to middle of terminal phalanx (webbing formula = I 0+–0+ II 0+–0+ III 0+–0+ IV 0+–0+ V); distinct, movable flap of skin on postaxial side of Toe V from middle of terminal phalanx to proximal end of metatarsus; distinct, movable flap of skin on preaxial side of Toe I from middle of terminal phalanx to level of inner metatarsal tubercle, continuing as weak fold on distal one third of tarsus; subarticular tubercles prominent, elevated, round; digits indicated by roman numeral (tubercle count in parentheses): V (2), IV (3) III (2), II (1), I (1); inner metatarsal tubercle oval, elongate with elevated post axial border.
Skin on top of head, throat, dorsal surfaces of forelimbs, and dorsum crenulate; skin on sides, around vent, and dorsal surfaces of thigh, shank and foot distinctly tuberculate, covered with moderately dense small, low glandular warts with translucent spinules; ventral skin smooth; pair of thin dorsolateral folds extend from post-orbital fold to mid-body, and although interrupted, on to the posterior third of body.
Color in preservative light brown dorsally and laterally; indistinct dark brown bars on upper lip, distinct and regularly spaced on lower lip; supratympanic fold dark brown; faint, dark brown crossbars on dorsal and lateral aspects of limbs, irregular dark brown spots on posterior thigh; throat mottled with light pigmentation, less pigmentation on mental patch; venter immaculate, ventral portions of limbs lightly dusted with melanophores; palmar and plantar surfaces dark brown, webbing between toes mottled.
Measurements. Morphometric data (in mm) for the holotype (male, FMNH 266636) are: SVL = 49.43; ED = 5.46; EN = 4.22; ES = 7.82; FEL = 25.48; FOL = 36.07; HL = 23.71; HW = 21.7; IN = 4.17; IO = 3.55; LAL = 10.27; MN = 21.54; PAL = 12.75; TBL = 25.54; UEW = 4.05; OL = 2.84; MD = 1.5.
Variation. Variation in body proportions given in Table 1 View TABLE 1 . All adult males have nuptial pads and spinule-covered mental patches. Relative head length (HL/SVL) is significantly larger (p = 0.02) in males (0.44–0.48) than in females (0.39–0.44). Based on the available data, it seems that females may attain a greater SVL, a trend contrary to the generally accepted paradigm of male-biased size dimorphism for the kuhlii Complex. A female-biased size dimorphism has been observed in other members of the kuhlii Complex (McLeod, pers. obs.), but it is possible that the observed variation in L. sisikdagu is the result of insufficient sampling.
Limnonectes kuhlii Limnonectes sisikdagu
Etymology. The specific name, sisikdagu , is derived from the Bhasa Indonesian words for “chin” (dagu) and “shield” (sisik), referring to the distinctive spinule-covered mental patch in males, a character that makes this frog unique among all the other known members of the kuhlii Complex. The specific name is used as a noun in apposition.
Comparisons. There are few obvious characters that differentiate one member of the kuhlii Complex from another. Fortunately, L. sisikdagu has a unique character—the presence of a spinule-covered mental patch in males— that distinguishes it from all other members of the kuhlii complex and from all other known members of the genus.
Limnonectes sisikdagu is a small member of the kuhlii Complex, and though the range in snout–vent length overlaps that of L. kuhlii from Java, the mean and maximum sizes attained by both males and females of L. sisikdagu are considerably smaller than those of L. kuhlii ( Table 1 View TABLE 1 ). Male L. kuhlii lack the nuptial pads that are present in L. sisikdagu . An externally visible tympanic annulus is present in some samples of L. kuhlii , but in none of the examples of L. sisikdagu . In dorsal view, the head of L. sisikdagu is more elongate and pointed than that of L. kuhlii (evidenced by the statisically significant difference in HL/HW ratios discussed below). All representatives of L. kuhlii examined here lack both the post-orbital fold and the dorsolateral fold present in L. sisikdagu . Though the tubercles do not extend as high onto the dorsum of the leg in L. kuhlii specimens, the tubercles are larger and more densely arranged than in L. sisikdagu ( Fig. 5 View FIGURE 5 ). These two species can also be distinguished by color in preservative, L. kuhlii being uniformly darker brown than L. sisikdagu .
Student's t-tests return significant differences among several mensural morphometric characters between the two species ( Table 2 View TABLE 2 ). Some characters differ significantly in only one sex, but when male/female data are considered together for the species, a significant difference between species can be observed (e.g., FEL/SVL ratio is different in females [p = 0.04] and when males and females are considered together [p = 0.02], but not when males are considered alone [p = 0.38]). Both sexes of L. sisikdagu have proportionately narrower heads (as determined by HL/HW) than either sex of L. kuhlii , in which head length and width are approximately equal. Additionally, the distance between eyes (as determined by IN/IO) is significantly narrower in L. sisikdagu than in either sex of L. kuhlii . Female L. sisikdagu have significantly smaller eyes (ED), larger eye–nares distances (EN), and shorter femurs (FEL) than do female L. kuhlii . Male L. sisikdagu have a greater upper eyelid width (UEW) relative to interorbital distance (IO) than do male L. kuhlii .
Student T-tests ( L. kuhlii : L. sisikdagu )
Distribution and ecology. Based on the specimens examined, Limnonectes sisikdagu occurs in central West Sumatra ( Fig. 6 View FIGURE 6 ). As with other members of the kuhii Complex, L. sisikdagu is found along forested streams. The new species seems to occur at elevations approximately between 520 and 1204 m. It is known to occur along streams in disturbed areas where forest has been cleared for agricultural exploitation.
The type locality, Batang Andaleh, is a tributary of Batang Tarsan, which is the main river from Solok and Lubuk Selasih to the coast. Main crops in this area are rice and tea, and the forest has been cleared for agriculture, mainly rice. Specimens from Batang Harau (near Harau Valley, Payakumbuh) were found at a river with protected forest border that varies from 50– 200 m. The lower story of forest is disturbed and includes bamboo, cinnamon, Melastoma, Dillenia, Dicranopteris, Hevea , and Uncaria . The river has clear water and a gravel bed with few boulders. Collections from Sarasah Bunta (near Harau Valley, Payakumbuh) were made along streams that flow from waterfalls. Batang Sarahah is a tributary of Batang Tarusan. The tree/shrub genera in this location include Gracinia, Elatostema, Rhinostaea, Eugenia, Zingiber, Calamus, and Areca .
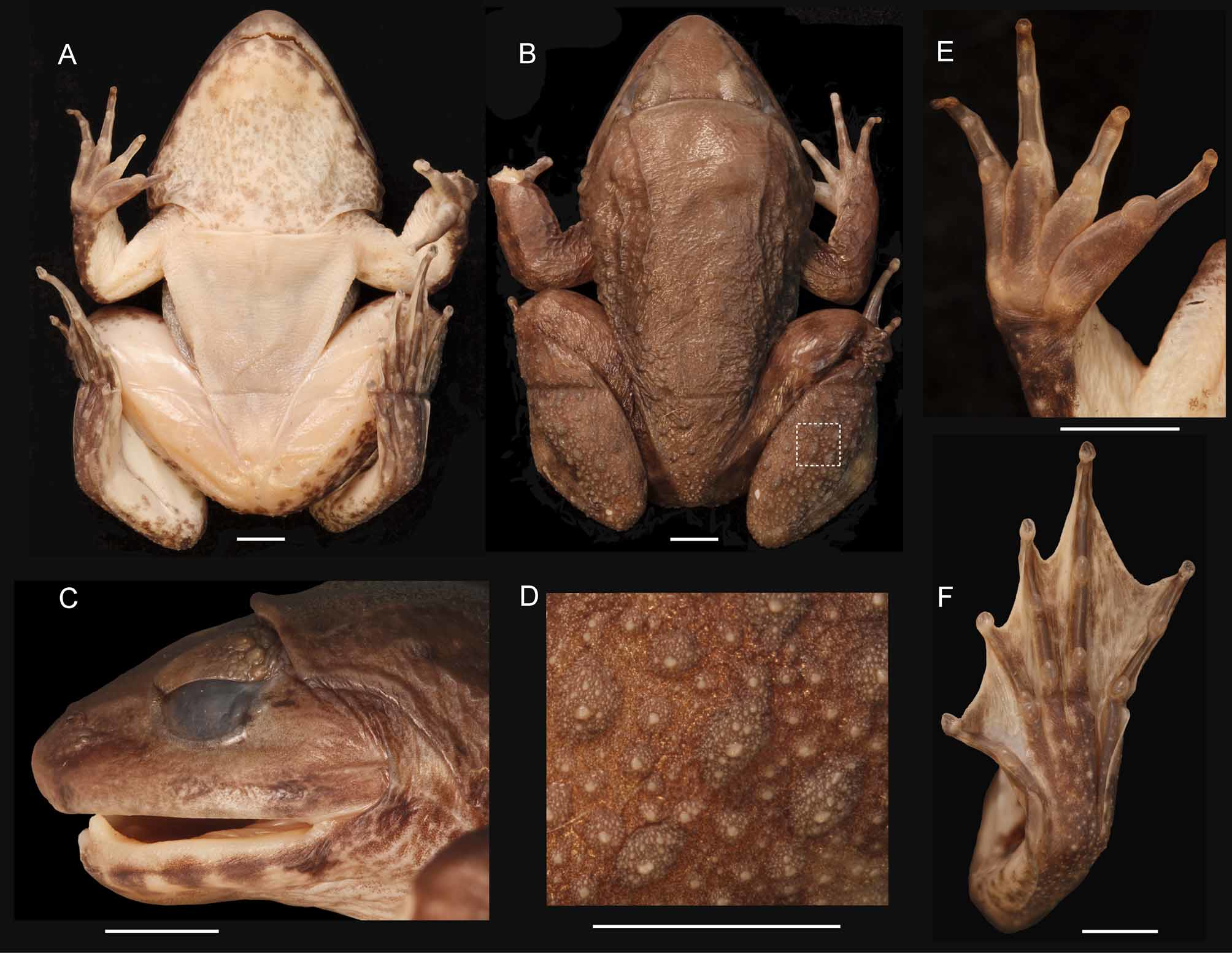




TABLE 1. Morphological measurements and ratios of specimens examined in this study. Ranges of values are presented for each character followed by the mean and standard deviation in parentheses.
| Character Male n = 4 | Female n = 13 | Male n = 5 | Female n = 4 |
|---|---|---|---|
| ED 4.71–9.17 (6.55 ± 1.92) | 5.41–8.15 (6.87 ± 0.86) | 4.32–5.46 (4.78 ± 0.45) | 4.76–5.40 (5.02 ± 0.27) |
| EN 3.25–7.06 (5.26 ± 1.58) | 3.35–5.71 (4.71 ± 0.71) | 3.28–4.32 (3.96 ± 0.42) | 3.54–4.80 (4.22 ± 0.55) |
| ES 5.14–12.14 (8.69 ± 2.87) | 5.84–10.54 (8.29 ± 1.4) | 5.92–7.82 (6.97 ± 0.73) | 6.88–7.81 (7.24 ± 0.42) |
| FEL 19.10–41.91 (31.64 ± 9.92) | 22.22–36.59 (30.41 ± 3.77) | 21.75–25.48 (22.28 ± 2.00) | 21.79–26.09 (23.89 ± 1.88) |
| FOL 27.19–48.85 (40.08 ± 9.86) | 30.84–43.38 (38.07 ± 3.96) | 28.91–36.07 (31.53 ± 2.89) | 30.76–38.66 (33.82 ± 3.79) |
| HL 15.28–40.25 (28.91 ± 10.42) | 17.55–31.28 (24.16 ± 3.71) | 18.53–23.71 (20.53 ± 2.22) | 17.99–22.33 (20.11 ± 1.83) |
| HW 15.24–40.8 (28.92 ± 10.62) | 16.43–29.55 (23.34 ± 3.47) | 16.25–21.70 (18.79 ± 2.28) | 16.06–20.67 (18.29 ± 1.95) |
| IN 3.43–6.68 (5.13 ± 1.33) | 3.81–5.80 (4.82 ± 0.56) | 3.24–4.17 (3.71 ± 0.34) | 3.30–4.05 (3.67 ± 0.31) |
| IO 2.76–7.97 (5.73 ± 2.2) | 3.13–5.86 (4.45 ± 0.77) | 2.31–3.55 (2.93 ± 0.57) | 2.48–3.03 (2.77 ± 0.25) |
| LAL 7.31–14.91 (11.74 ± 3.36) | 7.97–13.65 (10.52 ± 1.53) | 7.82–10.27 (8.62 ± 0.97) | 8.51–9.77 (9.15 ± 0.64) |
| MN 14.15–36.97 (26.08 ± 9.72) | 16.00–25.83 (20.76 ± 2.52) | 16.56–21.54 (18.32 ± 2.14) | 15.41–19.83 (17.42 ± 1.86) |
| PAL 9.79–19.36 (15.59 ± 4.54) | 11.45–17.98 (14.73 ± 1.65) | 9.87–12.75 (11.22 ± 1.18) | 11.14–12.46 (11.78 ± 0.59) |
| SVL 36.87–81.45 (62.39 ± 19.31) | 41.11–74.02 (59.41 ± 8.91) | 41.86–49.43 (45.36 ± 3.22) | 44.44–56.63 (49.10 ± 5.50) |
| RFL IV–V–II–III | IV–V–II–III | IV–V–III–II | IV–V–III–II |
| RTL IV–III–V–II–I | IV–III–V–II–I | IV–III–V–II–I | IV–III–V–II–I |
| TBL 18.40–36.26 (28.3 ± 7.89) | 20.49–31.52 (26.95 ± 2.87) | 19.80–25.54 (22.18 ± 2.24) | 21.03–26.34 (23.46 ± 2.44) |
| TYE 0–5.76 (3.17 ± 2.92) UEW 2.87–6.18 (4.52 ± 1.35) | 3.63–5.50 (4.52 ± 0.58) 2.97–6.23 (4.97 ± 0.85) | NA 3.02–4.05 (3.55 ± 0.40) | NA 3.05–4.03 (3.56 ± 0.45) |
| OL 1.49–7.65(4.14 ± 2.6) | 1.60–3.68 (2.62 ± 0.6) | 1.52–3.16 (2.38 ± 0.64) | 1.69–2.35 (1.91 ± 0.30) |
| MD 0.86–3.19 (1.87 ± 0.97) | 1.05–1.86 (1.42 ± 0.25) | 0.99–1.50 (1.21 ± 0.19) | 0.99–1.19 (1.07 ± 0.09) |
| PAL/LAL 1.28–1.42 | 1.22–1.67 | 1.23–1.45 | 1.24–1.34 |
| PAL/SVL 0.23–0.27 | 0.23–0.28 | 0.24–0.26 | 0.21–0.25 |
| OL–MD/HL 0.04–0.11 | 0.02–0.08 | 0.03–0.09 | 0.03–0.05 |
| ED/HL 0.2–0.31 | 0.24–0.33 | 0.22–0.24 | 0.23–0.27 |
| HL/HW 0.99–1.01 | 0.96–1.08 | 1.07–1.14 | 1.08–1.12 |
| HL/SVL 0.41–0.49 | 0.38–0.43 | 0.44–0.48 | 0.39–0.43 |
| SVL–HL/ 0.51–0.59 SVL | 0.57–0.62 | 0.52–0.56 | 0.57–0.61 |
| IN/IO 0.80–1.24 | 0.91–1.31 | 1.09–1.60 | 1.27–1.35 |
| IN/SVL 0.07–0.09 | 0.06–0.09 | 0.08–0.08 | 0.07–0.08 |
| EN/IN 0.95–1.10 | 0.84–1.21 | 0.89–1.33 | 0.99–1.38 |
| UEW/IO 0.68–1.04 | 0.92–1.37 | 1.08–1.52 | 1.14–1.54 |
| EN/ES 0.58–0.63 | 0.47–0.62 | 0.54–0.62 | 0.51–0.66 |
| TBL/SVL 0.44–0.50 | 0.42–0.5 | 0.47–0.52 | 0.47–0.49 |
| FEL/SVL 0.49–0.52 | 0.49–0.54 | 0.47–0.52 | 0.46–0.50 |
| LAL/SVL 0.18–0.20 | 0.15–0.21 | 0.17–0.21 | 0.17–0.20 |
TABLE 2. Selected results from inter-specific comparisons of morphological characters using 2 - tailed Student’s t-tests (p values).
| Character ED/HL | Male 0.836 | Female 0.030 | Male+Female 0.018 |
|---|---|---|---|
| HW/HL | 0.000 | 0.005 | 0.000 |
| IN/IO EN/IN UEW/IO | 0.037 0.518 0.010 | 0.002 0.016 0.075 | 0.001 0.019 0.014 |
| TBL/SVL | 0.132 | 0.132 | 0.011 |
| FEL/SVL | 0.148 | 0.026 | 0.003 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |