
Hymenocephalus lethonemus Jordan & Gilbert
|
publication ID |
https://doi.org/ 10.11646/megataxa.3.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/B711B23F-FF07-86C6-D99D-C791FE7679F6 |
|
treatment provided by |
Plazi |
|
scientific name |
Hymenocephalus lethonemus Jordan & Gilbert |
| status |
|
Hymenocephalus lethonemus Jordan & Gilbert View in CoL in Jordan & Starks, 1904
[Japanese name: Itodara]
( Figs. 137–138 View FIGURE 137 View FIGURE 138 ; Appendix 3-8B)
Hymenocephalus lethonemus View in CoL Jordan & Gilbert in Jordan & Starks, 1904:615, unnumbered fig. [original description; holotype: USNM 50936, from Sagami Bay, Albatross sta. 3697, in 120–265 ftm (220–485 m); paratypes from Tokyo market and Sagami Bay]; Jordan et al. 1913:413, fig. 383 (listed; Japan; new Japanese name: “Ito-dara”); Gilbert & Hubbs 1916:188 (brief description; 22 spec. from East China Sea, off Cape Shionomisaki, Suruga Bay, and Sagami-nada); Okada & Matsubara 1938:452 (in key; Japan); Matsubara 1955:1315 (in key; Japan); Kamohara 1964:96 (listed; Kochi Pref.); Matsubara 1965:507 (compiled; Japan); Okamura 1970a:54, pl. II, text-fig. 23 (description; biological notes; 83 spec. from Pacific off southern Japan from Sagami Bay to Kochi and East China Sea); Okamura 1970b: table 1 (listed; Japan); Kuroda 1971:56 [listed from Suruga Bay after Okamura (1970a)]; Tominaga & Uyeno 1981:489 (listed; Japan); Yatou 1982:143, 347, fig. 87 [brief description; 7 spec. from Kyushu-Palau Ridge and Tosa Bay; photo based on BSKU spec. (catalog no. unknown)]; Ohta 1983: table A (listed; Suruga Bay); Okamura 1984a:201, 358, fig. 142 (brief description; 13 spec. from Okinawa Trough; photo based on BSKU 29790); Okamura 1984b:92, pl. 80, fig. G (compiled); Okamura 1988:92, pl. 80, fig. G (compiled); Nakabo 1993:355 (in key; Japan); Shinohara et al. 1996:169 (25 spec. listed from Pacific off Tohoku); Okamura 1997:128, fig. 2 (compiled); Shinohara & Matsuura 1997:291 (listed; Suruga Bay); Nakabo 2000:419 (in key; Japan); Shinohara et al. 2001:305 (19 spec. listed from Tosa Bay); Nakabo 2002:419 (in key; Japan); Yoda et al. 2002:11 (listed; East China and Yellow Seas); Chiou et al. 2004a:44, fig. 12, table 1 (brief description; 13 spec. from Da-xi; first record from Taiwan); Shinohara et al. 2005:417 (1 spec. listed from Ryukyu Islands); Suetsugu & Ohta 2005: tables 2–3 (listed; southeast of Boso Peninsula and Enshunada); Senou et al. 2006:421 (listed; Sagami Sea); Shinohara & Williams 2006:551, fig. 2K (listed; Sagami Sea); Kitagawa et al. 2008:39, unnumbered fig. (brief description; spec. from Pacific off Tohoku); Shao et al. 2008b: table 2 (21 spec. listed from northeastern and southwestern Taiwan, and South China Sea); Shinohara et al. 2009:708 (listed; Pacific off Tohoku); Motomura et al. 2010:77, fig. 34 (5 spec. listed from Yakushima Island, Kagoshima Pref.); Furuhashi et al. 2010: table 2 (5 spec. listed from northern Okinawa Trough); Nakabo & Kai 2013:495 (in key; Japan); Schwarzhans 2014:42, fig. 18 (description; comparison; in key); Iwamoto et al. 2015:75 (brief description; 36 spec. from northeastern and southwestern Taiwan, and Philippines); Motomura 2020:39 (listed; Japan).
Hymenocephalus striatissimus View in CoL (not Jordan & Gilbert in Jordan & Starks 1904): Okamura 1997:128, fig. 3 [in part, photo spec. (BSKU 41914)].
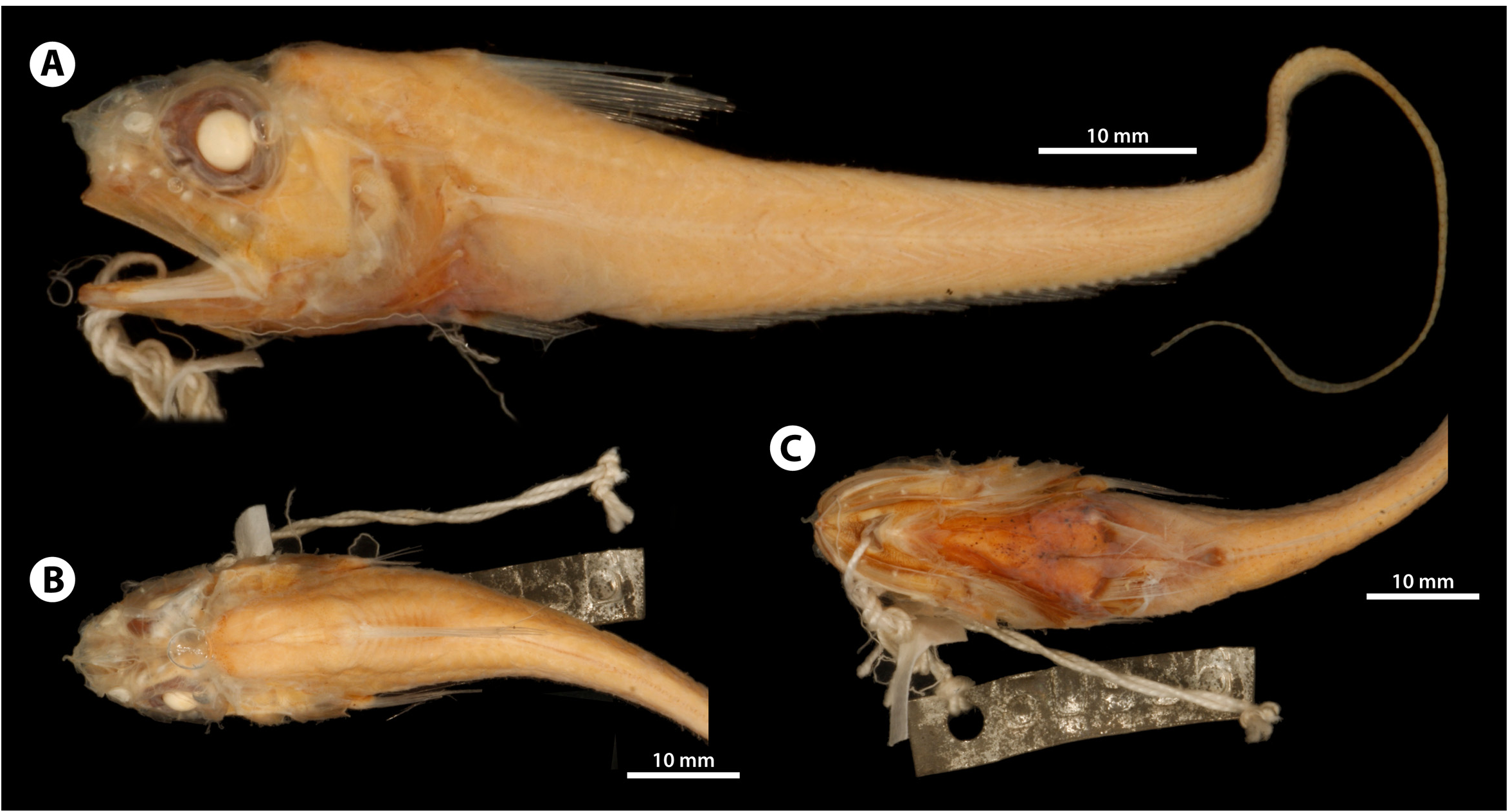
Diagnosis. A species of Hymenocephalus with 11 (rarely 12) pelvic-fin rays; chin barbel absent or rudimentary, length less than 1% PRL; snout high, not depressed, length 37–44% PRL; orbit diameter 43–46% PRL; interorbital width 38–41% PRL; orbit–preopercle distance 55–60% PRL; suborbital width 16–17% PRL; first dorsal-fin rays II,11; pectoral-fin rays i14–i17; gill rakers on first arch (outer/inner) 17–20/24–26, on second arch 24–26/22–25; numerous tiny black melanophores densely and uniformly scattered over trunk and tail, giving dusky appearance to body; caudal vertebrae barely visible externally; ostial and caudal colliculi of otolith not fused.
Material examined. 8 specimens. Holotype of Hymenocephalus lethonemus: USNM 50936 (25.9 mm HL, 140 mm TL), off Manazuru , Sagami Bay, Japan, 35.1333ºN, 139.1667ºE, 120–265 ftm (220–485 m), Albatross sta. 3697, 8-ft Tanner beam trawl, 5 Mar. 1900 GoogleMaps . Paratypes of H. lethonemus: USNM 51455 (3, 15.7–23.9 mm HL, 86+–126+ mm TL, counts and measurements taken on 1 of 3), off Numazu , Suruga Bay, 35.0333ºN, 138.7833ºE, 63–75 ftm (116–138 m), Albatross sta. 3707, 8-ft Tanner beam trawl, 8 Mar. 1900 GoogleMaps . Non-type : Japan : BSKU 109012 View Materials (1, 27.2 mm HL, 112+ mm TL), off Amami , East China Sea, 28.5513ºN, 127.0375ºE, 621 m, T/ V Nagasaki-maru, cr. N365, sta. J6, tr. 3, 3-m ORE beam trawl, coll. N. Nakayama, 16 Nov. 2012 GoogleMaps ; * BSKU 44762 View Materials (1, 30.7 mm HL, 155+ mm TL), Tosa Bay , 600 m, FRV Kotaka-maru, otter trawl, 16 May 1988 ; BSKU 13348 View Materials (1, 30.4 mm HL, 169+ mm TL), west of Muroto , Tosa Bay, 350–450 m, 14 May 1968 ; BSKU 44766 View Materials (1, 26.5 mm HL, 153+ mm TL), Tosa Bay , 600 m, FRV Kotaka-maru, otter trawl, 16 May 1988 .
Counts and measurements. Based on 5 specimens (23.9–30.4 mm HL, 112+–169+ mm TL). Counts: first dorsal-fin rays II,11; pectoral-fin rays i14–i17; pelvic-fin rays 11; gill rakers on first arch (outer/inner) 17–20/24– 26, on second arch 24–26/22–25; longitudinal scales 17–19; transverse scale rows below first dorsal-fin origin 4.5–5, below first dorsal-fin midbase 2–2.5, below second dorsal-fin origin 3–3.5.
The following measurements are in% of HL, followed by those in % of PRL in parentheses: snout length 28–32 (37–44); orbit diameter 32–35 (43–46); postorbital length 45–46 (60–63); postrostral length 74–76; orbit–preopercle distance 41–45 (55–60); suborbital width 12–13 (16–17); upper-jaw length 51–54 (68–71); length of rictus 44–45 (59–60); length of premaxillary tooth band 39 (52–53); preoral length 14–17 (19–23); distance between tip and lateral angle of snout 18–22 (23–30); snout width 25–30 (33–41); internasal width 19–24 (26–32); interorbital width 28–31 (38–41); occipital width 13–14 (17–19); body width over pectoral-fin bases 41–55 (54–73); body depth at first dorsal-fin origin 66–68 (89–92); body depth at anal-fin origin 41–49 (54–66); prepelvic length 100–121 (132–162); preanus length 151–158 (199–212); preanal length 152–161 (200–215); isthmus–pelvic distance 43– 57 (58–77); isthmus–anus distance 87 (117); isthmus–anal distance 89–109 (120–143); pelvic–anal distance 46–58 (60–76); pelvic-fin length 82–92 (110–120); pectoral-fin length 55–63 (73–84); predorsal length 96–105 (129– 143); height of first dorsal fin 74–81 (97–109); length of first dorsal-fin base 30–36 (40–47); interdorsal length 59–66 (78–90); length of gill slit 30–33 (39–45); length of posterior nostril 4–7 (5–9); barbel length 0–1 (0–1).
Size. To about 19 cm TL ( BSKU 33501 View Materials , 185 View Materials + mm TL, Okinawa Trough , Japan).
Development. Early life stage was briefly described by Endo (2014), based on three specimens (7.5–9.5 mm HL) collected from Sagami Bay.
Distribution. Restricted to Japan and Taiwan. Known from the Pacific off Japan northward to the Oshika Peninsula (38.32ºN), Okinawa Trough, northern Taiwan, Izu Islands, and Kyushu-Palau Ridge, at depths of 116‾ 1000 m ( Chiou et al. 2004a; Shao et al. 2008a, 2008b; Schwarzhans 2014; Iwamoto et al. 2015; Appendix 3-8B). This species is well represented on the upper continental slope along the Pacific coasts of Japan, and is one of the numerically dominant grenadiers in Tosa Bay, Kumano-nada, and off the Boso Peninsula ( Suetsugu & Ohta 2005; Horikawa 2009: appendix 3).
Remarks. For further morphological information see the original description ( Jordan & Gilbert in Jordan & Starks 1904) and Okamura (1970a); otolith morphology was detailed by Schwarzhans (2014).
Hymenocephalus lethonemus was described on the basis of specimens collected from Sagami Bay (type locality) and “ Tokyo market”, Japan ( Fig. 138 View FIGURE 138 ). Although Weber (1913) reported H. lethonemus from Indonesian waters, Gilbert & Hubbs (1920:535) considered Weber’s specimens to be representatives of their new species, H. nascens , described from the Philippines and East Indies. Gilbert & Hubbs (1920) distinguished these taxa solely by modal counts of pelvic-fin rays [usually 12 (very rarely 11 or 13) in H. nascens vs. usually 11 (12) in H. lethonemus ]. Subsequently, Weber & de Beaufort (1929:61) regarded nascens as a subspecies of H. lethonemus , noting that “we are no more able to discover any other difference and feel therefore not authorized to separate the specimens from the Archipelago [viz., nascens ; note by the present author] specifically from those from Japan, known as H. lethonemus ”. While all subsequent authors recognized each taxon as a full species (e.g., Okamura 1970a; Marshall & Iwamoto 1973; Iwamoto 1990; Iwamoto & Merrett 1997; Iwamoto & Williams 1999; Schwarzhans 2014; Iwamoto et al. 2015), further morphological studies are required to elucidate their taxonomic status, particularly based on well-preserved materials.
Relationships and comparisons. Hymenocephalus lethonemus belongs to the H. striatulus group as defined by Schwarzhans (2014). In addition to H. lethonemus , this group includes: H. billsam Marshall & Iwamoto, 1973 disjunctly distributed in the northwestern Atlantic and off Brazil; H. nascens widespread in the East Indies and Australasian waters; and H. striatulus Gilbert, 1905 known only from Hawaii and the Sala y Gomez Ridge. According to Schwarzhans (2014), H. lethonemus substantially differs from the above three species in that its pelvic-fin rays usually number 11 [rarely 12 vs. 12– 15 (rarely 11 in H. nascens )]. It further differs from all except H. nascens in its absence of the chin barbel (vs. a small but distinct barbel present in H. billsam and H. striatulus ). The reader refers to Schwarzhans (2014) for further comparisons of the H. striatulus group.
Due to their close resemblance, H. lethonemus may be confused with H. papyraceus Jordan & Gilbert in Jordan & Starks, 1904 in Japanese waters. They share the absence of the chin barbel and similar pigmentation pattern consisting of small black melanophores densely scattered over the trunk and tail. However, H. lethonemus readily differs from H. papyraceus in having more gill rakers (24–26 vs. 18–22 on the outer side of the second arch; 22–25 vs. 15–21 on the inner side of the second arch). It further differs from H. papyraceus in its narrower suborbital space (16–17% PRL vs. 22–26%), shorter pectoral fin (73–84% PRL vs. 95%), longer pelvic fin (110–120% vs. 79%), and shorter, lower, and narrower snout (preoral length 19–23% PRL vs. 28–39%; snout width 33–41% PRL vs. 44–53%; internasal width 26– 32% PRL vs. 34–39%).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |