
Smeringopina bineti (Millot, 1941)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3713.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C5F0BC11-92C0-4B30-9DB3-200882AC8950 |
|
DOI |
https://doi.org/10.5281/zenodo.6161969 |
|
persistent identifier |
https://treatment.plazi.org/id/B20287ED-FFEF-FF96-B990-C561FA6C39F0 |
|
treatment provided by |
Plazi |
|
scientific name |
Smeringopina bineti (Millot, 1941) |
| status |
|
Smeringopina bineti (Millot, 1941) View in CoL
Figs. 2–3 View FIGURES 2 – 16 , 41–45 View FIGURES 34 – 45 , 54 View FIGURES 46 – 55 , 76–95 View FIGURES 76 – 82 View FIGURES 83 – 95
Smeringopus bineti Millot 1941: 24 –27, figs. 10A–G, 10bis A–F.
Smeringopina bineti: Huber 1995: 299 (transfer to Smeringopina ). Dimitrov, Astrin & Huber 2013 (DNA data).
Types. ♂ lectotype (designated herein) from Guinea, Dalaba [10°40.7’N, 12°15.7’W], and 1♂ 7♀ 5 juvs. paralectotypes from Guinea, Dalaba (3♀), Kindia [10°00.8’N, 12°48.6’W] (1♀), Grotte de Tinka [10°44.1’N, 12°16.3’W] (1♀) and Grotte de Tassacouré (locality not identified; 1♂ 2♀ 3 juvs), all collected by J. Millot in 1937, all together in one vial, original label: “ Smeringopus bineti, Millot rec. et det., Afrique occidentale franc.”, in MNHN (Ar 10490), examined.
Notes. Since the numbers of adult and juvenile specimens in the single vial agrees well with the total number of specimens studied by Millot (1941), I assume that all type specimens from all four localities were joined.
A lectotype is designated because specimens from Koumbaya and Grotte de Tassacouré differ slightly in their morphology (see below), as already noted by Millot (1941). The larger male (from Dalaba, illustrated in Millot’s figure 10F) is chosen because the locality of the other male (Grotte de Tassacouré) could not be identified. The male lectotype is in bad condition (see below) but Millot’s figure leaves no doubt that it is conspecific with new material collected at the type locality and at other places in Guinea.
Other material examined. GUINEA: Moyenne-Guinée: Dalaba (10°40.7’N, 12°15.7’W), along brook in forest, 920 m a.s.l., 21.xi.2008 (B.A. Huber), 4♂ 3♀ in ZFMK (Ar 10176); same data, 1♂ 4♀ 2 juvs. in pure ethanol, in ZFMK (Gui 108); same locality at 10°40.6’N, 12°16.0’W, 1150 m a.s.l., 21.xi.2008 (B.A. Huber), 1♂ 6♀ 2 juvs. in pure ethanol, in ZFMK (Gui 101); Dalaba, forest fragment at 10°41.5’N, 12°15.9’W, 1230 m a.s.l., 21.xi.2008 (B.A. Huber), 1♀ in pure ethanol, in ZFMK (Gui 116); near Dalaba, Pont de Dieu (10°41.1’N, 12°12.8’W), sheltered rock cavities, 1150 m a.s.l., 21.xi.2008 (B.A. Huber), 1♀ 7 juvs. in pure ethanol, in ZFMK (Gui 107). Near Maréla (10°09.9’N, 11°30.4’W), road cut, 420 m a.s.l., 26.xi.2008 (B.A. Huber), 3♂ 8♀ 1 juv. in ZFMK (Ar 10177); near Maréla (10°09.8’N, 11°17.1’W), 630 m a.s.l., 4.xii.2008 (B.A. Huber), 1♂ in ZFMK (Ar 10178); same data, 1♂ 2♀ 2 juvs. in pure ethanol, in ZFMK (Gui 89). SE Mamou (10°17.6’N, 11°56.5’W), forest along brook, 460 m a.s.l., 4.xii.2008 (B.A. Huber), 7♂ 11♀ 1 juv. (2 vials) in ZFMK (Ar 10179-80); same data, 2♂ 2♀ in pure ethanol, in ZFMK (Gui 88). Near Sêbori, in cave (10°46.6’N, 12°17.4’W), 1010 m a.s.l., 22.xi.2008 (B.A. Huber), 4♂ 15♀ (2 vials) in ZFMK (Ar 10181-82); same data, 3♂ 2♀ in pure ethanol, in ZFMK (Gui 99). Near Doucki, canyon (10°59.6’N, 12°35.3’W), 1020 m a.s.l., 24.xi.2008 (B.A. Huber), 4♂ 2♀ in ZFMK (Ar 10183); same data, 2♀ 4 juvs. in pure ethanol, in ZFMK (Gui 79). Basse-Guinée: near Koumbaya (10°10.3’N, 12°53.3’W), forest, 220 m a.s.l., 19.xi.2008 (B.A. Huber), 2♂ 4♀ in ZFMK (Ar 10184); same data, 2♀ 1 juv. in pure ethanol, in ZFMK (Gui 94). Near Kindia (10°00.8’N, 12°48.6’W), along brook in plantation forest, 540 m a.s.l., 5.xii.2008 (B.A. Huber), 1♀ in pure ethanol, in ZFMK (Gui 98).
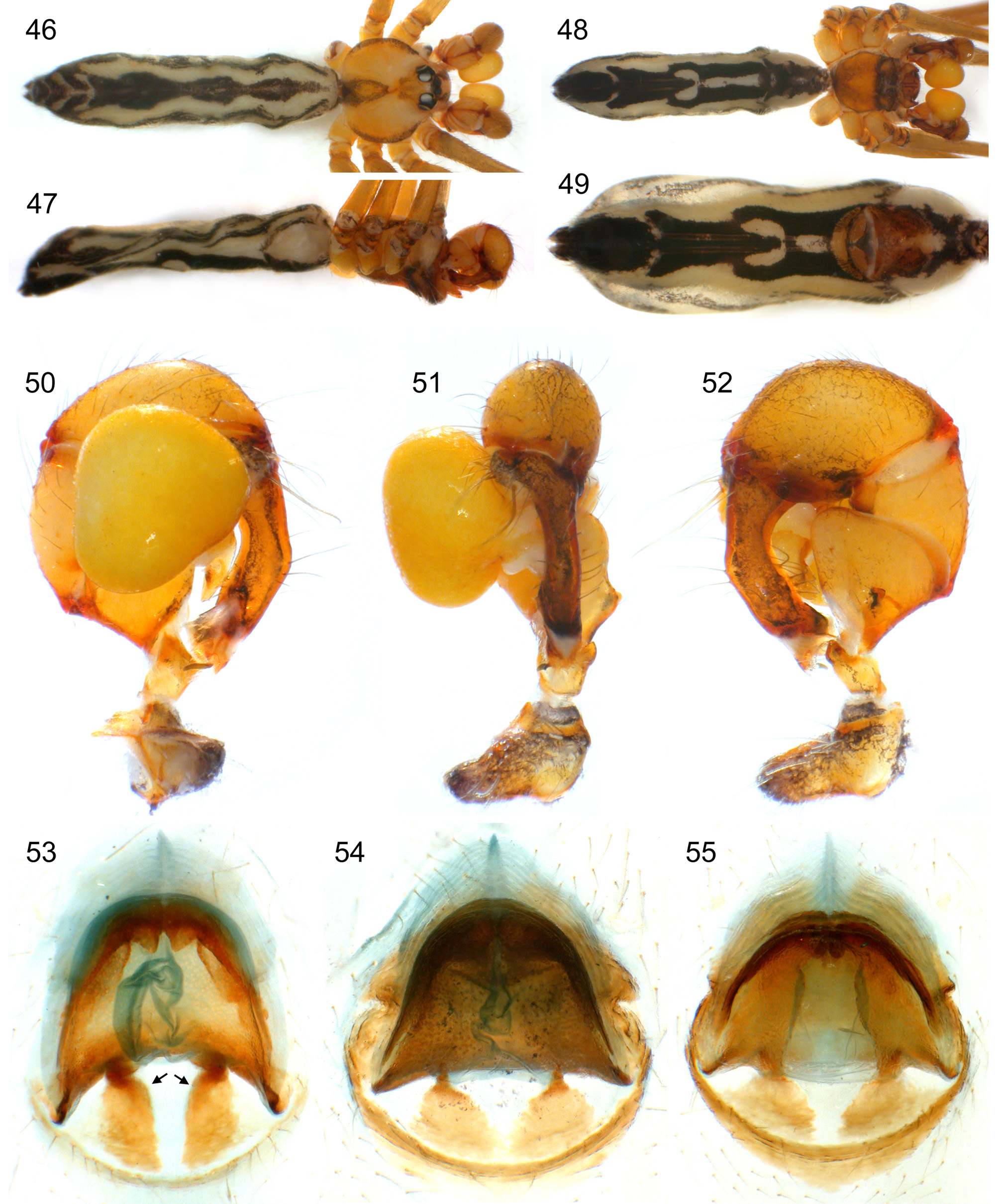
Diagnosis. Distinguished from similar congeners ( S. guineensis , S. pulchra ) by shorter abdomen, shorter legs, and dark lateral marks on carapace (in addition to dark margin; Fig. 41 View FIGURES 34 – 45 ); from S. guineensis also by presence of ventro-distal apophysis on procursus ( Fig. 77 View FIGURES 76 – 82 ), all modified hairs on large cheliceral apophyses grouped close together ( Fig. 79 View FIGURES 76 – 82 ), and low hump on anterior epigynal plate; from S. pulchra also by pair of median projections proximally on male chelicerae ( Fig. 83 View FIGURES 83 – 95 ), curved procursus ( Figs. 45 View FIGURES 34 – 45 , 76–77 View FIGURES 76 – 82 ), pointed bulbal apophysis ( Fig. 78 View FIGURES 76 – 82 ), and only weakly curved posterior margin of anterior epigynal plate ( Figs. 42 View FIGURES 34 – 45 , 80 View FIGURES 76 – 82 ).
Male (near Maréla). Total body length 6.0, carapace width 1.7. Leg 1: 48.9 (11.7 + 0.6 + 11.6 + 22.3 + 2.7), tibia 2: 7.8, tibia 3: 5.4, tibia 4: 7.6; tibia 1 L/d: 73. Distance PME-PME 170 µm, diameter PME 150 µm, distance PME-ALE 60 µm, distance AME-AME 45 µm, diameter AME 125 µm. Carapace ochre-orange with brown triangular mark posteriorly, brown lateral margins and indistinct lateral marks; ocular area brown, clypeus with pair of brown lines and brown rim, sternum red-brown; legs ochre, tips of femora and tibiae whitish, dark rings subdistally on femora and tibiae and in patella area; abdomen ochre-gray with distinct black pattern dorsally, laterally, and ventrally. Habitus as in Fig. 41 View FIGURES 34 – 45 , ocular area slightly elevated, secondary eyes with indistinct ‘pseudolenses’ ( Fig. 87 View FIGURES 83 – 95 ); clypeus unmodified except sclerotized rim, rim roughly straight; deep thoracic pit and pair of shallow furrows diverging behind pit. Chelicerae as in Fig. 79 View FIGURES 76 – 82 , with small median projections proximally ( Fig. 83 View FIGURES 83 – 95 ), lateral proximal apophyses, large distal apophyses provided with 3–4 modified hairs each, and very small distal apophyses close to fang joints. Palps as in Figs. 43–45 View FIGURES 34 – 45 ; coxa with prominent sclerotized rim retrolaterally; trochanter with simple retrolatero-ventral apophysis; femur with small retrolateral apophysis pointing dorsodistally, with large ventral bulge distally, without ventral or prolateral modification proximally; prolateral femurpatella joint only slightly moved toward ventrally; tarsus with about four stronger hairs; procursus without hinge, curved toward ventrally, with two long ventral hairs bent around procursus and directed toward dorsally, distally with distinctive ventral apophysis and moveable dorsal sclerite embedded in membranous cuticle ( Figs. 76–77 View FIGURES 76 – 82 , 84– 85 View FIGURES 83 – 95 ); bulb with simple pointed apophysis with small subdistal branch, arising from membranous basal projection ( Fig. 78 View FIGURES 76 – 82 ; sperm duct apparently opens at membranous basal projection at basis of pointed apophysis). Legs without spines and curved hairs, with few vertical hairs, retrolateral trichobothrium on tibia 1 at 1%; prolateral trichobothrium present on all tibiae; pseudosegments barely visible. ALS with eight spigots each ( Fig. 88–89 View FIGURES 83 – 95 ); gonopore with two epiandrous spigots ( Fig. 90 View FIGURES 83 – 95 ).
Variation. Number of modified hairs on male cheliceral apophyses slightly variable, even within one specimen. Males from Koumbaya and Grotte de Tassacouré with slightly different ventral apophysis distally on procursus (basis of apophysis simply rounded rather than complex as in Fig. 77 View FIGURES 76 – 82 ; cf. fig. 10E in Millot 1941) and with smaller moveable dorsal sclerite ( Fig. 86 View FIGURES 83 – 95 ). Tibia 1 in 19 other males: 9.1–12.6 (mean 11.3); small cheliceral apophyses near fang joints in some specimens barely visible; sternum variably dark. The type specimens are bleached but mostly in fair condition, many legs missing, palps and chelicerae missing in lectotype, right palp missing in paralectotype.
Female. In general similar to male; sternum variably dark as in males, sometimes almost black. Tibia 1 in 39 females: 8.9–10.5 (mean 9.7). Epigynum consisting of large anterior plate with low hump in median part and large posterior plate composed of sclerotized posterior arc and pair of distinct anterior plates ( Fig. 42 View FIGURES 34 – 45 ); internal genitalia as in Figs. 54 View FIGURES 46 – 55 and 82 View FIGURES 76 – 82 , with distinct globular structures between uterus externus and anterior epigynal plate ( Fig. 81 View FIGURES 76 – 82 ). Epigynal shape apparently slightly variable (cf. figs. 10bis D–F in Millot 1941) but most of this variation is due to variable pigmentation. ALS as in male.
Natural history. S. bineti was found both in well preserved forests (e.g. under rocks along a stream near Dalaba) and in degraded patches (e.g. in small holes of roadcut near Maréla). It was also common in Sêbory cave, where it occupied small cavities in the cave wall. It vibrated rapidly when disturbed and was seen to build both domed and spherical webs near Koumbaya.
Distribution. Known from several localities in western Guinea (Moyenne-Guinée and Basse-Guinée, Fig. 33 View FIGURE 33 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |