
Verruca punica Buckeridge & Jagt
|
publication ID |
https://doi.org/ 10.5281/zenodo.183257 |
|
DOI |
https://doi.org/10.5281/zenodo.6230520 |
|
persistent identifier |
https://treatment.plazi.org/id/AA7BBC04-8F22-685F-FF6F-F95FFB63FBBB |
|
treatment provided by |
Plazi |
|
scientific name |
Verruca punica Buckeridge & Jagt |
| status |
sp. nov. |
Verruca punica Buckeridge & Jagt , sp. nov.
(Figs 3A–G; 4A–R)
Type material. Holotype NHMM 2007 0 94, a specimen lacking moveable scutum and moveable tergum; basis unknown; height 1.1 mm, length 2 mm, width 1.2 mm. Paratypes NHMM 2007 0 95, 2007 0 96, 2007 0 97 and 2007 100, and NMV P314992.
Material examined. In addition to the types (see above), sample AFN 601 yielded four moveable terga, one moveable scutum, a single fixed scutum, one fixed tergum, a single carina plus>20 incomplete plates ( NHMM 2007 0 98, 2007 0 99, 2007 101 and 2007 102a), all handpicked by one of us (RPS) in the Spring of 1988 from washed microfossil residues of the AFN sample set collected in 1982 by a crew of the Cretaceous- Palaeogene Boundary Working Group at El Haria, some 5 km southwest of El Kef ( Smit et al., 1997).
Diagnosis. Moderately small Verruca , elongate, with a relatively smooth shell and large pentangular orifice; moveable tergum quadrangular, carinal margin slightly longer than basal margin, with prominent apicobasal rib extending beyond the basal margin, and two secondary articular ribs, upper rib upwardly inflected; moveable scutum with a prominent, apico-basal rib, and a secondary rib on the upper margin.
Description. Shell small, slightly compressed laterally; apex of carina produced so that carina lies beneath the operculum that is subparallel to the base (Fig. 3C); longitudinal ribbing very weak to imperceptible but with clearly formed transverse growth striae; orifice large, pentangular; rostrum broad, comprising approximately one third of shell wall, two ribs interlocking with carina, one with a quadrangular fixed scutum; fixed tergum narrow (width less than one third height); moveable tergum subrhomboidal, first (= upper) articular rib raised, rounded and slightly produced at scutal margin; second articular rib slightly depressed, closer to apico-basal (primary) rib than to upper, protrudent with other ribs to produce sinuous scutal margin; apico-basal rib rounded in section, elevated, gently arcuate, concave on upper margin, widening towards base, where it extends beyond basal margin by a distance approximating basal width; upper carinal angle c. 90°; carinal-basal angle c. 110°, upper scutal angle c. 80°; internal surface with slight apical depression for adductor muscle attachment, otherwise weakly undulating. Moveable scutum triangular, almost twice as long as wide, exterior with a sharply raised apico-articular ridge which has a beaded appearance due to intersection with transverse growth ridges; secondary rib on movable scutum on a gently concave tergal margin; basal margin very slightly convex, occludent margin broadly arcuate; apical angle c. 50°; basi-rostral angle c. 90°, basi-tergal angle c. 120°; external apico-basal striae on both the moveable tergum and scutum is either absent or very weak.
Etymology. In allusion to the Punic wars (264–241 BC, 218–201 BC and 149–146 BC), in which Rome crushed Carthaginian power, and obliterated Carthage, on the coast of northern Tunisia.
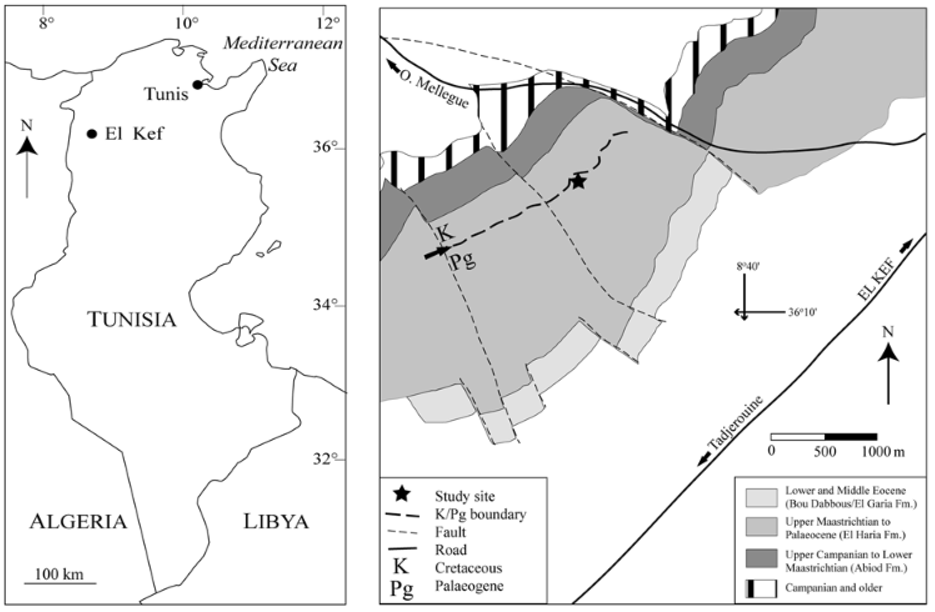
Geographic and stratigraphic setting. Sample AFN 601 was taken 15 m above the K/Pg boundary in the so-called KS section at El Kef (co-ordinates: 36N08'57''/08E38'39''; see Fig. 1 View FIGURE 1 ). This is within the El Haria Formation, which straddles the Cretaceous-Palaeogene boundary and comprises shales and marls deposited in the Tunisian Trough ( Smit et al., 1997; Molina et al., 2006). About 13 m above sample AFN 601 a set of four white, marly limestones interrupt the clay-marl sequence and form a subtle pale ridge along the main SW-NE aligned valley. Microfossil assemblages (planktic and benthic foraminifera, dinoflagellate cysts, and ostracods) from the interval comprising the cirripede material studied here document an open marine, oligotrophic environment and deposition at outer neritic to bathyal depths (~ 200 m) during the early Palaeocene ( Kouwenhoven et al., 1997; Guasti et al., 2005). There was no macrofauna associated with sample AFN 601, but it does come from a 5–10 m interval with common and unusually large (up to 2 mm) agglutinated foraminifera.
Biostratigraphically, the sample is allocated to the lower part of calcareous nannofossil zone NP2 and planktic foraminiferal zone P1b ( Guasti et al., 2005). This level correlates with the middle of Chron 29N and has an age of ~64.9 Ma ( Luterbacher et al., 2004) to ~64.4 Ma ( Berggren & Pearson, 2005).
FIGURE 3A–G. Verruca punica Buckeridge & Jagt , sp. nov. from the Lower Palaeocene (Danian) El Haria Formation, El Kef area (northwest Tunisia). A, basal view; B, top view, showing pentangular-shaped opercular opening; C, detail of B, showing rostrum; D, lateral view, made up of rostrum, fixed scutum and fixed tergum; E, rostral view; F, carinal view; G, lateral view, made up of carina and rostrum (left to right). All views are of specimen NHMM 2007 0 94 (holotype).
Remarks. The above description of the shell, as being “slightly compressed laterally”, is based upon a single, almost complete specimen (the holotype). The wall plates in this specimen show no evidence of deformation (e.g. stress cracks) and conform to the few disarticulated wall plates available for study. However, it is possible that further collecting may show the shell to have generally been more circular in plan; because of this, this character is not included in the diagnosis. The moveable tergum and scutum possess no clearly preserved apico-basal striae, however it is most likely that these would be present; their absence is likely due to modification (partial dissolution) of any fine ornamentation on calcareous remains during diagenesis.
The first true verrucomorphs were probably very much like the modern relic taxon Neoverruca brachylepadoformis Newman , in Newman & Hessler, 1989, which possesses a five-plated wall and confirms a monophyletic origin of the balanomorph barnacles from the Scalpelliformes ( Newman & Hessler, 1989; Buckeridge & Newman, 2006). However, Neoverruca differs from Verruca sensu stricto by the possession of a medial latus interposed between the fixed tergum and fixed scutum, and the presence of several whorls of imbricating plates that elevate the wall above the substratum, thus having the appearance of a scalpelliform. Verruca punica Buckeridge & Jagt , sp. nov. is clearly a verrucid, and although there are some very minor fractures at the base of the shell wall, these are considered artifacts of preservation rather than the remains of any imbricating plate.
The description provided here is based upon the assumption that all the recovered material that is ‘verrucid’ is from the same species. Unfortunately, the only ‘near-complete’ specimen lacks opercula, and there are no other complete carina or rostra in the sample. There are, nonetheless, some excellent examples of moveable terga and some moveable scuta, and association of these could fit the opercular opening of the single shell. Unfortunately, this single shell is slightly smaller than would be required to accommodate most of the opercula recovered. However, size difference alone is not seen as an impediment to this association. If further material is located, and is able to demonstrate that there is more than one taxon, then the holotype, without opercula, will need a revised diagnosis. The approach adopted here is thus parsimonious, and in reality is the norm rather than the exception in much of palaeontology.
FIGURE 4. Verrucomorph and scalpellimorph cirripede plates from the Lower Palaeocene (Danian) El Haria Formation, El Kef area (northeast Tunisia); A–R. Verruca punica Buckeridge & Jagt , sp. nov.; A, B, moveable tergum (NHMM 2007 0 95, paratype), exterior and interior aspects; C, D, moveable tergum (NHMM 2007 0 96, paratype), exterior and interior aspects; E, F, moveable scutum (NHMM 2007 0 97, paratype), exterior and interior aspects; G, H, moveable tergum (NMV P 314992, paratype), exterior and interior aspects; I, J, incomplete moveable tergum (NHMM 2007 098), exterior and interior aspects; K, L, incomplete moveable tergum (NHMM 2007 099), exterior and interior aspects; M, N, fixed tergum (NHMM 2007 100, paratype), exterior and interior aspects; O, P, incomplete moveable tergum (NHMM 2007 101), exterior and interior aspects; Q, R, incomplete moveable tergum (NHMM 2007 102a), exterior and interior aspects; S, T, Scillaelepas sp.?, incomplete carina (NHMM 2007 102b), exterior and interior aspects; U, V, Scillaelepas sp.?, lateral plate (NHMM 2007 102c), exterior and interior aspects; W, X, Scillaelepas sp.?, lateral plate (NHMM 2007 103), exterior and interior aspects; Y, Z, Scillaelepas sp.?, rostrolateral plate (NHMM 2007 104), exterior and interior aspects.
There are also fragments of other cirripedes present (Fig. 4S–Z), and although these are incomplete, they are not verrucid. Rather they can be interpreted as scalpelliform, and likely belong either to Calantica Gray, 1825 , or Scillaelepas Seguenza, 1876 .
Verruca punica Buckeridge & Jagt , sp. nov. may be distinguished from other Late Cretaceous and early Palaeocene verrucids by the absence of longitudinal ribbing on the exterior and a laterally compressed, rather than subcircular shell (Fig. 3A). In particular, the rather produced carina differentiates it from all other species of Ve r r u c a and in this character it shows similarity to Altiverruca View in CoL and Metaverruca View in CoL . However, it is distinguished from the former (see e.g., Young, 1998) by having an operculum that is subparallel to the basis, and from the latter by the elongate shape, simpler wall plates (i.e., fewer interlocking ribs) and shape of the operculum, which is D-shaped in Metaverruca View in CoL (see Foster & Buckeridge, 1995; Buckeridge, 1997). The Pliocene Verruca alaskana also has a produced plate in the shell wall, resulting in an elongated, rather than rounded shell, but in that species, the produced plate is the rostrum. Unlike most extant species of Ve rr u ca, the shell of V. punica Buckeridge & Jagt , sp. nov. is imperforate. An impunctate shell is a character that is apparently shared with the fossil taxa V. t. tasmanica , V. t. chatheca , V. nuciformis , V. prisca , V. pusilla and V. rocana , although perforations are present in the Miocene V. digitali ( Buckeridge & Finger, 2001) and the Pliocene V. koikei ( Koike et al., 2006) .
Although verrucid remains have been recovered as disarticulated plates, especially from friable coquinas (where they are easily separated from other shells; e.g., Verruca tasmanica tasmanica and V. tasmanica chatheca ), most fossil verrucids are found articulated and attached to shelly substrates. In part this is an artifact of collecting, as verrucid fragments are small and often overlooked. Of the few that are known from fine-grained sediments similar to those at El Kef, Verruca digitali is of note: it is locally very abundant, and occurs as sheets along with associated bryozoans in a shale. As there are no obvious molluscs or other suitable hardshell substrates at the horizon, the verrucids (and bryozoans) are interpreted to have grown on marine algae and/or seagrass leaves, of which there are now no remains ( Buckeridge & Finger, 2001).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SuperOrder |
Thoracica |
|
Order |
|
|
Family |
|
|
Genus |
Verruca punica Buckeridge & Jagt
| Buckeridge, John S., Jagt, John W. M. & Speijer, Robert P. 2008 |
V. koikei (
| Koike et al. 2006 |
V. digitali (
| Buckeridge & Finger 2001 |