
Lamellomorpha australis Kelly & Cárdenas, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.506 |
|
publication LSID |
lsid:zoobank.org:pub:0D5F8DFB-C1AC-47F5-9129-C9241DF3DB04 |
|
DOI |
https://doi.org/10.5281/zenodo.5619280 |
|
persistent identifier |
https://treatment.plazi.org/id/2A7459B3-0FBA-441B-806C-7C988075843A |
|
taxon LSID |
lsid:zoobank.org:act:2A7459B3-0FBA-441B-806C-7C988075843A |
|
treatment provided by |
Plazi |
|
scientific name |
Lamellomorpha australis Kelly & Cárdenas |
| status |
sp. nov. |
Lamellomorpha australis Kelly & Cárdenas View in CoL sp. nov.
urn:lsid:zoobank.org:act:2A7459B3-0FBA-441B-806C-7C988075843A
Figs 1 View Fig , 3 View Fig , 6 View Fig ; Tables 1–2 View Table 1 View Table 2
Lamellomorpha strongylata View in CoL — Bergquist 1968: 31 –32, pl. 11f (in part). — Dumdei et al. 1997: 2636 – 2639. — Li et al. 1998: 724 –728. — Hickford 2007: 29 –41. — Blunt & Munro 2008: 285, 1798, 1854. — Buckingham et al. 2010: 349. — Bycroft & Payne 2013: 429, 1642.
Etymology
Named for the Chatham Rise and Subantarctic New Zealand distribution of this species (‘ australis ’, south, Latin).
Type material
Holotype
NEW ZEALAND • Subantarctic region of New Zealand, Bounty Platform, NZOI Station A 751; 47.743° S, 179.124° E; depth 155 m; 16 Nov. 1962; NIWA 89736 View Materials leg.; Agassiz medium trawl; UPSZTY 178605 (a small piece of the holotype preserved in 70% ethanol, as well as a spicule preparation), NIWA. GoogleMaps
Paratypes
NEW ZEALAND – same collection data as for the holotype; NIWA 92896 View Materials to 92900 View Materials , 93483 View Materials , 93484 View Materials , and 93487 leg.; NIWA GoogleMaps • same collection data as for preceding; NIWA 93485 View Materials leg.; UPSZTY 178606 (a small piece of the paratype preserved in 70% ethanol), NIWA GoogleMaps • same collection data as for preceding; NIWA 93486 View Materials leg.; UPSZTY 178607 (a small piece of the paratype preserved in 70% ethanol), NIWA GoogleMaps .
Type locality
Subantarctic region of New Zealand, Bounty Platform; depth 155 m.
Other material examined
NEW ZEALAND • Bounty Platform , NZOI Station I 711; 47.833° S, 179.250° E; depth 139 m; 22 Mar. 1979; NIWA 89717 View Materials leg.; rock dredge; NIWA GoogleMaps • Bounty Platform , NZOI Station A714; 47.725° S, 179.067° E; depth 165 m; 5 Nov. 1962; NIWA 86733 View Materials leg.; cone and mesh dredge; NIWA GoogleMaps • Bounty Platform , NZOI Station A715; 47.683° S, 179.051° E; depth 121 m; 5 Nov. 1962; NIWA 89720 View Materials leg.; cone and mesh dredge; NIWA GoogleMaps • Bounty Platform , NZOI Station A757; 47.693° S, 179.058° E; 17 Nov. 1962; NIWA 113894 View Materials leg.; NIWA GoogleMaps • Solander Trough , NZOI Station D39; 50.967° S, 165.750° E; depth 549 m; 7 May 1963; NIWA 44388 View Materials leg.; gear dredge, cone mesh with bag; NIWA GoogleMaps • Campbell Platform , NZOI Station B184; 52.615° S, 169.117° E, depth 344 m; 11 Oct. 1959; NIWA 93499 View Materials leg.; dredge; NIWA GoogleMaps • Macquarie Ridge , NIWA Station TAN0803/69; 52.398° S, 160.657° E; depth 451– 438 m; 9 Apr. 2008; NIWA 40328 View Materials leg.; epibenthic sled; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 427; 43.077° S, 175.272° E; depth 180–237 m; 20 Feb 1995; NIWA 44240 View Materials leg.; Agassiz Trawl; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 446; 43.245° S, 175.444° E; depth 71–76 m; 22 Feb 1995; NIWA 44261 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 447; 43.245° S, 175.458° E; depth 80–85 m; 22 Feb 1995; NIWA 44263 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 448; 43.240° S, 175.458° E; depth 74 m; 22 Feb 1995; NIWA 44267 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 446; 43.247° S, 175.444° E; depth 71–76 m; 22 Feb 1995; NIWA 44272 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 435; 43.172° S, 175.340° E; depth 108–113 m; 20 Feb 1995; NIWA 137202 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , North Mernoo Bank, NIWA Station W 446; 43.247° S, 175.444° E; depth 71–76 m; 22 Feb 1995; NIWA 137201 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , South Mernoo Bank, NIWA Station W 452; 43.450° S, 175.135° E; depth 120–180 m; 22 Feb 1995; NIWA 44241 View Materials leg.; rock dredge; NIWA GoogleMaps • Chatham Rise , South Mernoo Bank, NIWA Station W 454; 43.451° S, 175.109° E; depth 126–130 m; 22 Feb 1995; NIWA 44262 View Materials leg.; rock dredge; NIWA GoogleMaps .
Description
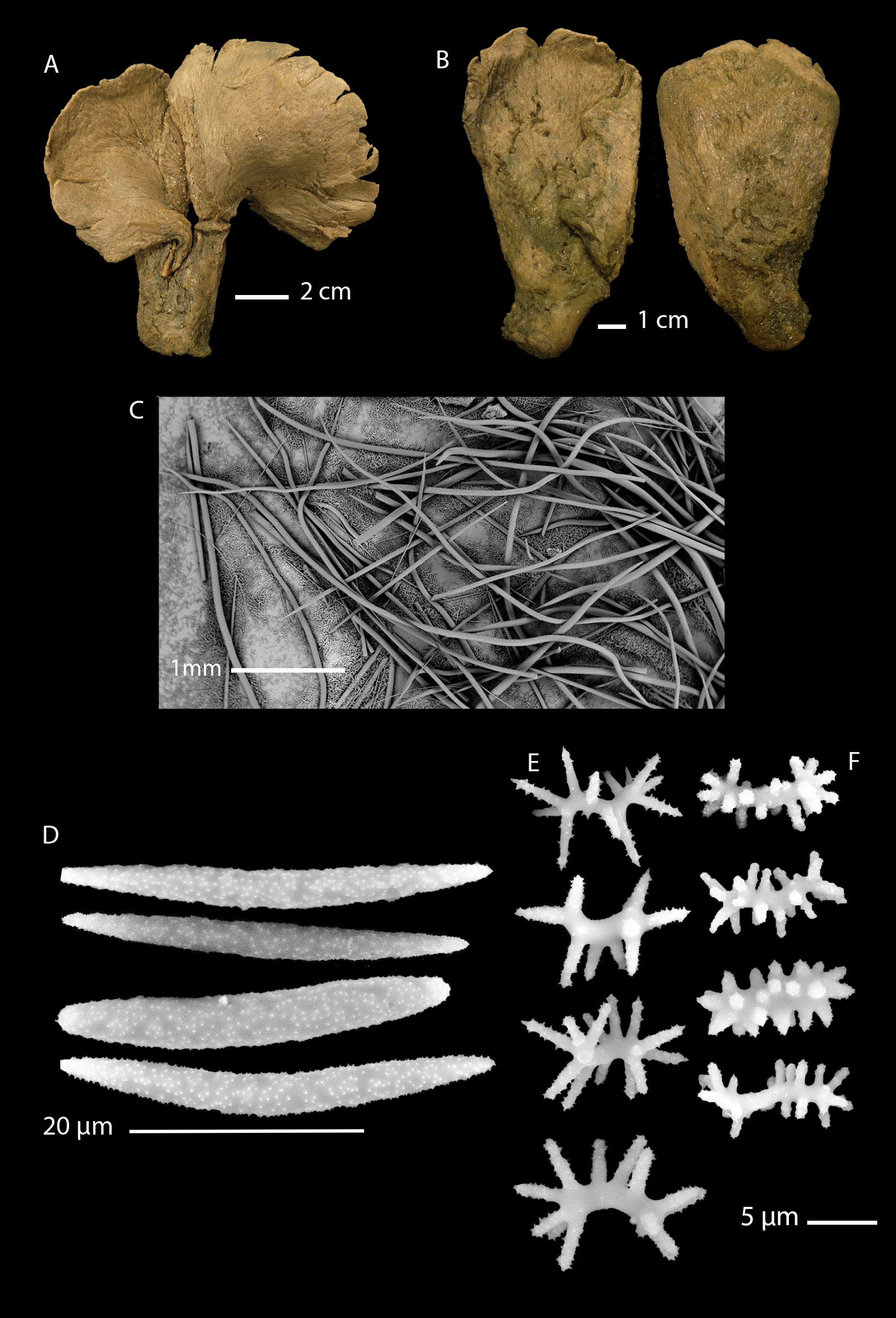
Uni- to bilamellate fan sponge, table tennis bat-shaped ( Fig. 3A View Fig ) or club-shaped ( Fig. 3B View Fig ), 130–200 mm high with a short, broad stalk, 2–3 cm thick, and a relatively thick lamella (up to 2 cm thick in places), attenuating towards the margins, which are frequently incised. The specimen from NZOI Station B176 was described by Bergquist (1968) as being 160 mm high, 89 mm wide, and 32–58 mm thick, supported by a stalk that was broken and thus was not measured. Oscules were not visible in the holotype or any other specimen. Pores are inconspicuous and compressed (probably due to the fixation) and were 40– 80 µm in diameter (measured on the holotype). Surface relatively smooth with low ridges radiating from the stalk to the fan margins. Texture, relatively soft, compressible. Colour in life and preservative, tan.
Skeleton
Choanosome disorganised, with megascleres orientated more or less parallel with the axis of the fan and stalk ( Fig. 6 View Fig B–D), with single or a couple of megascleres extending beyond the surface from the subectosome. The ectosome is extremely thick and packed with microstrongyles and streptasters, which also occur in great density throughout the sponge.
Spicules
MEGASCLERES ( Table 1 View Table 1 ; Fig. 3C View Fig )
Bergquist (1968) considered the megascleres of the “subantarctic specimen” (presumably the NZOI Station B 176 specimen from Campbell Plateau) to have, “predominantly oxeas, some of which are curved, but most are contort”. Our examination of new material reveals that oxeas dominate the megasclere complement; these are rarely to never modified; all have sharp attenuated tips. The majority are straight to slightly curved and contort, but not to the degree seen in L. strongylata . The megascleres reach their greatest length in L. australis sp. nov., up to 3575 µm long in the specimen NIWA 93486 leg. (paratype).
MICROSCLERES ( Table 2 View Table 2 ; Fig. 3 View Fig D–F)
Bergquist did not differentiate between the microscleres of the holotype of L. strongylata (from the Three Kings) and the subantarctic Campbell Plateau specimens ( L. australis sp. nov.), calling them all “plesiasters” in the table of spicule dimensions. However, in pl. 11, figs E2 and F2–3, a clear difference is obvious between the illustrations of the streptasters: they are metasters in pl. 11, fig. E2 ( L. strongylata ) and larger metasters (pl. 11, fig. F3) and “abnormal microrhabds” in pl. 11, fig. F2 ( L. australis sp. nov.). The “abnormal microrhabds” of Bergquist (1968: pl. 11, fig. F2) are most likely spirasters (as in our Fig. 3F View Fig ), the ornamentation of which would not have been visible under light microscopy available at the time.
Thus, L. australis sp. nov. has three forms of microsclere: a microxea ( Fig. 3D View Fig ) with attenuating, hastate ends that is usually straight, but may be slightly curved, ranging in length from about 19–40 µm; metaster-like streptasters with heavily spined, relatively long rays ( Fig. 3E View Fig ), ranging in length from about 7–15 µm; spirasters with abundant, short, microspined rays that emanate from a long, spiral axis ( Fig. 3F View Fig ), ranging in length from about 8–14 µm.
Distribution
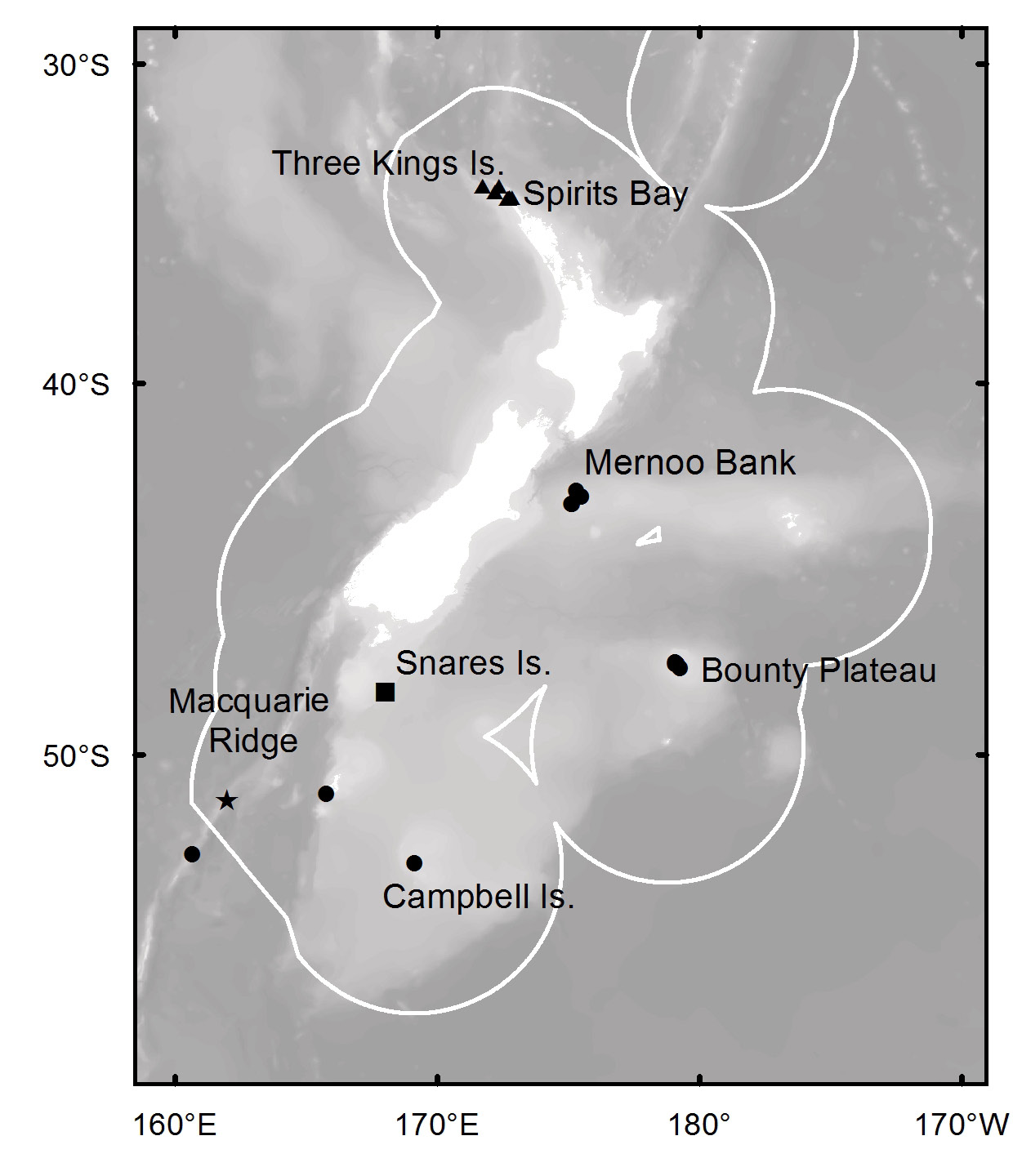
Subantarctic region of New Zealand: Mernoo Bank (depth 71–237 m), north-western Chatham Rise ( Dumdei et al. 1997; Li et al. 1998); Bounty Platform (depth 121–165 m), Solander Trough (depth 549 m), Campbell Platform (depth 344 m), and Macquarie Ridge (depth 451– 438 m).
Substrate, depth range and ecology
Attached by a thick stalk to sediment covered rocky substrate, depth 71– 549 m.
DNA barcodes
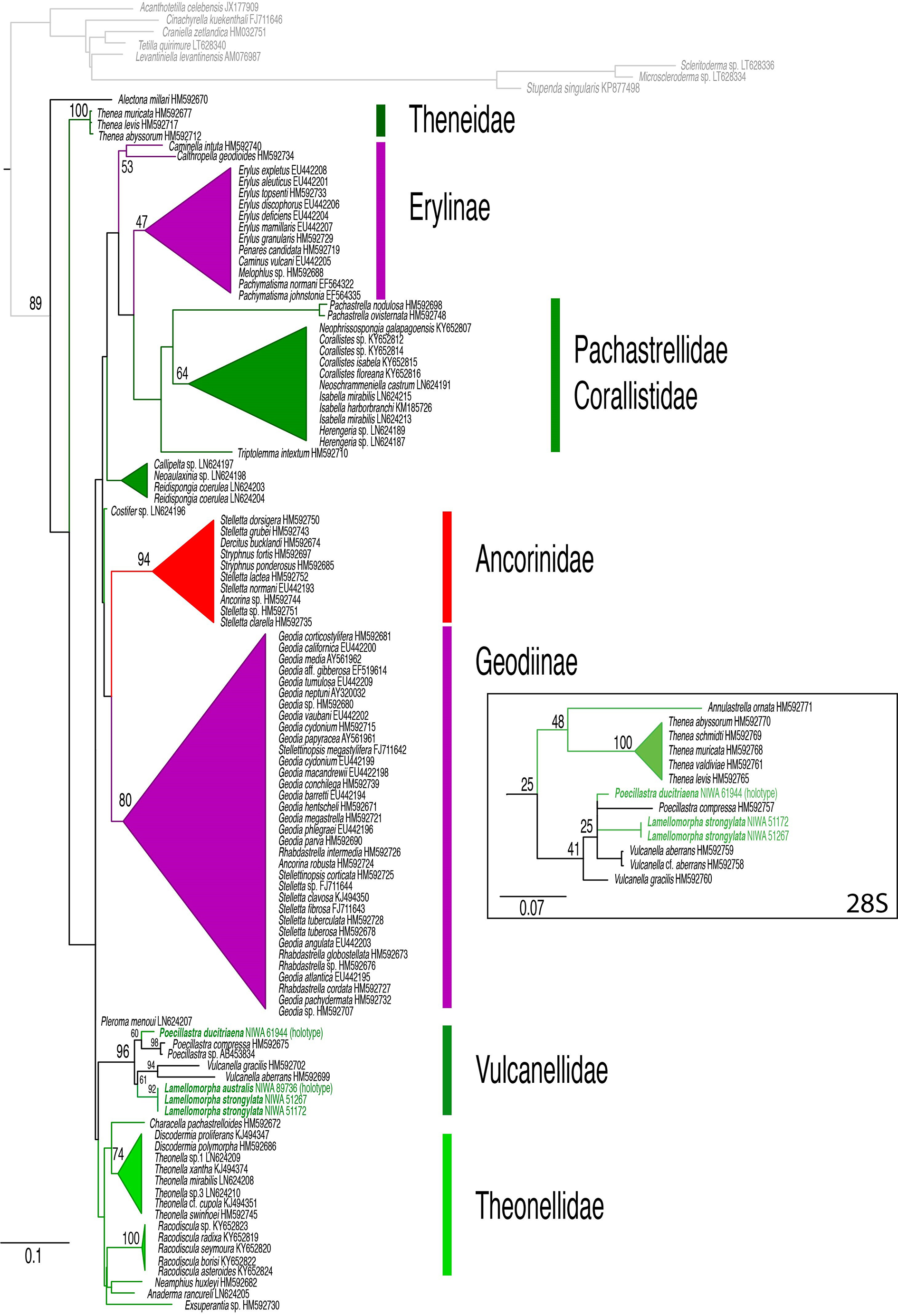
COI. Holotype (minibarcode, MK 033625 View Materials ), no bp differences with the COI minibarcode of L. strongylata .
Remarks
Bergquist (1968) listed two specimens from Campbell Plateau in the Subantarctic region of New Zealand, from NZOI Station B176 (46 fathoms = 84.12 m) and NZOI Station B184 (103 fathoms = 188.4 m). Unfortunately, neither specimen was found in the NIWA collections and both are presumed lost. However, we did find a specimen from NZOI Station B184 (NIWA 93499 leg.) from a depth of 344 m. Bergquist (1968) considered the two specimens she examined to be conspecific with L. strongylata , despite the obvious disjunct distribution, but noted that the subantarctic specimens had predominantly oxeas, an observation we agree with.
Examination of numerous specimens uncovered in NIC allows us to convincingly separate L. australis sp. nov. from the type species on geographic distribution, morphology, and skeletal details, despite the COI minibarcodes not differentiating them ( Fig. 7 View Fig ). The most obvious difference that separates L. australis sp. nov. from L. strongylata is the markedly disjunct distribution and depth ranges: L. strongylata has only been recorded to the north of New Zealand, 41–200 m depth, while L. australis sp. nov. is only found on and south of Mernoo Bank on the Chatham Rise, ranging in depth from 71 m on Mernoo Bank, to 549 m in the Solander Trough. In terms of morphology and colouration in life, L. strongylata forms a relatively soft, dark royal blue, palmate sponge, supported by a relatively narrow stalk, while L. australis sp. nov. forms a distinctive, tan, paddle-shaped sponge, with thin, incised margins, on a thick, short stalk. In terms of skeletal architecture, the choanosome of L. australis sp. nov. is much more densely packed with microscleres than L. strongylata , and the former species lacks the relatively distinct subectosomal tracts of the latter. As noted by Bergquist (1968), the megascleres of L. australis sp. nov. differ from those of L. strongylata in being predominantly oxeas (straight and contort) with no modifications of the tips to strongyloxeas as in L. strongylata . In addition, we note that the contort forms are much longer on average, and have a greater size range, than in L. strongylata . Finally, microscleres also discriminate L. australis sp. nov. from L. strongylata . Lamellomorpha strongylata has stubby, often centrotylote roughened microstrongyle, while L. australis sp. nov. has a relatively fine, curved, slightly longer roughened microxea. Lamellomorpha strongylata has metaster- to amphiaster-like streptasters with heavily spined, relatively long rays in one size category, while L. australis sp. nov. has metaster-like streptasters with heavily spined, relatively long rays and spirasters with abundant, short, microspined rays that emanate from a long, spiral axis. Spirasters are absent in L. strongylata .
As part of their ongoing investigations into New Zealand marine natural products in sponges, professors Murray Munro and John Blunt and their group in the Department of Chemistry, University of Canterbury, Christchurch, collected what was identified by the late professor Patricia Bergquist as L. strongylata , from Mernoo Bank on the Chatham Rise. Vouchers of these sponge specimens were donated to NIC for their preservation and future study and have been re-identified here as L. australis sp. nov., extending the known distribution of L. australis sp. nov. north to the Chatham Rise. Thus, it is L. australis sp. nov., and not L. strongylata , from which biologically active secondary metabolites were isolated by the University of Canterbury group, including calyculins (A, B, E, and F), calyculinamides (A and B), swinholide H ( Dumdei et al. 1997), and theonellapeptolides ( Li et al. 1998; Hickford 2007); identical and related compounds are found in sponges in the genus Theonella Gray, 1868 and Discodermia du Bocage, 1869 (family Theonellidae ).
It has been shown that calyculins and its derivatives (e.g., calyculinamides) could be produced by the filamentous bacteria ‘ Entotheonella ’ spp. in Discodermia ( Wakimoto et al. 2014) . ‘ Entotheonella ’ spp. are especially abundant in Theonella swinhoei , as well as in many other demosponges ( Wilson et al. 2014). In her PhD thesis, Hickford (2007) noticed that filamentous heterotrophic (Gram positive) bacteria were very abundant in L. australis sp. nov. and were associated with several theonellapeptolides. The producer of theonellapeptolides is currently unknown but the results of Hickford (2007) suggest that L. australis sp. nov. may be a host for theonellapeptolides-producing ‘ Entotheonella ’-like bacteria. Hickford (2007) also isolated unicellular bacteria from the same specimens and showed these were associated with swinholide H. This result concurs with previous results from Bewley et al. (1996), who identified swinholide A in unicellular bacteria isolates from Theonella swinhoei from Palau. However, it is an apparent contradiction with Ueoka et al. (2015) who convincingly show that misakinolide A (swinholide-like compound) from another Theonella swinhoei chemotype (chemotype WA from Japan) is produced by ‘ Entotheonella serta ’. Therefore it seems that swinholide-type compounds may be produced by bacteria other than ‘ Entotheonella ’ in L. australis sp. nov. and Theonella swinhoei (chemotype Palau). Hickford (2007) further states that specimens from the “northern population of L. strongylata ” ( L. strongylata ) not only had very low quantities of filamentous bacteria (apparently limited to the surface of the sponge), but also missed the biological activity, and therefore may not produce the above mentioned compounds. Thus, to conclude, L. strongylata and L. australis sp. nov. also clearly differ in terms of natural products and microbial communities.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Astrophorina |
|
Family |
|
|
Genus |
Lamellomorpha australis Kelly & Cárdenas
| Kelly, Michelle, Cárdenas, Paco, Rush, Nicola, Sim-Smith, Carina, Macpherson, Diana, Page, Mike & Bell, Lori J. 2019 |
Lamellomorpha strongylata
| Buckingham J. & Baggaley K. H. & Roberts A. D. & Szabo L. F. 2010: 349 |
| Blunt J. W. & Munro M. H. G. 2008: 285 |
| Hickford S. J. H. 2007: 29 |
| Li S. & Dumdei E. J. & Blunt J. W. & Munro M. H. & Robinson W. T. & Pannell L. K. 1998: 724 |
| Dumdei E. J. & Blunt J. & Munro M. H. G. & Pannell L. K. 1997: 2636 |
| Bergquist P. R. 1968: 31 |