
Ophryotrocha diadema Åkesson, 1976
|
publication ID |
https://doi.org/ 10.5281/zenodo.202108 |
|
DOI |
https://doi.org/10.5281/zenodo.5619578 |
|
persistent identifier |
https://treatment.plazi.org/id/9D5787CE-FFCD-0355-FF40-3725E67A40DB |
|
treatment provided by |
Plazi |
|
scientific name |
Ophryotrocha diadema Åkesson, 1976 |
| status |
|
Ophryotrocha diadema Åkesson, 1976 View in CoL
Figure 1 View FIGURE 1 A; 2; 3A–C; 8; 9; 10; 11; Table 1 View TABLE 1 ; 2
Ophryotrocha diadema Åkesson, 1976: 23 View in CoL , figs 1–6; Åkesson & Costlow 1978; Åkesson 1982; Pleijel & Eide, 1996; Dahlgren et al. 2001; Heggøy et al. 2007; Simonini et al. 2009, 2010; Macnaughton et al. 2010.
Material examined. Live cultures from specimens collected in a small-boat marina, Long Beach, California, U.S.A., by Bertil Åkesson, 1989. Additional preserved material from same culture: 50 specimens ( SMNH 117237), 0.5–2.1 mm long (5–18 chaetigers); 50 specimens (AM W.37221), 0.5–2.3 mm long (4–18 chaetigers).
Description. Length of most live adults 2.5–3.5 mm (15–17 chaetigers), maximum length 4.6 mm (25 chaetigers) ( Table 1 View TABLE 1 ). Live animals ( Fig. 1 View FIGURE 1 A) translucent, preserved opaque white. Prostomium anteriorly rounded, wider than long, dorsoventrally flattened, bearing pair of dorsal cylindrical antennae, palps absent, replaced by ciliated pads ( Figs. 2 View FIGURE 2 A, B). Anterior part of prostomium and tips of antennae with small bundles of adjoined tactile cilia, appearing spike-like in living animals. Ciliary bands separated by gaps; two bands of cilia encircling prostomium, one complete circle in front of antennae, and one incomplete circle at level of antennae. Latter interrupted by antennae; cilia between antennae slanting posteriorly, separated by small mid-dorsal space, giving appearance of eyebrows. Additional ventral semicircle of cilia anterior to complete circle. Posterior border of prostomium indicated dorsally by four nuchal organs, ventrally by mouth, lateral restrictions almost absent.
Peristomium represented by two apodous, achaetous segment-like rings, each surrounded by complete circle of cilia. Two eyes, slightly below border between prostomium and peristomium, between inner and outer nuchal organs. Eyes internal, visible only in living animals. Eyes with refractive crystalline platelets as typical for Ophryotrocha , appearing silvery white under incident light. Crystalline platelets irregularly branching, extending medially and anteriorly, forming triangular luminous area together with eyes ( Fig. 1 View FIGURE 1 A). Following chaetigers with dorsal and ventral ciliary bands, laterally interrupted by parapodia.
Parapodia uniramous, with acicular lobe and ventral retractile lobe supported by short simple chaeta; dorsal and ventral cirri absent ( Fig. 2 View FIGURE 2 C, D). Supra-acicular fascicle consisting of 1–3 simple chaetae, subacicular fascicle with 2–3 heterogomph falcigers and inferiormost simple chaeta ( Fig. 2 View FIGURE 2 E). Appendages of falcigers and upper part of simple chaetae serrated with simple distal tip ( Fig. 2 View FIGURE 2 F). Mammillate rosette glands, two per segment, present dorsolaterally on up to 5 posterior segments ( Fig. 2 View FIGURE 2 C, D, G). Pygidium bearing dorsal anus and pair of dorsal pygidial cirri; dorsal stylus absent ( Fig. 2 View FIGURE 2 C).
Jaws consisting of mandibles and maxillae; former permanent, latter undergoing at least two moults. Mandibles consisting of two elongate shafts, widening distally into bifid serrated cutting plates with 22–28 tiny teeth and large blunt sublateral tooth-like projection at anterior cutting edge of each plate ( Fig. 3 View FIGURE 3 A). Shafts of adult animals 110–170 µm long, with basal and distal lateral thickenings ( Fig. 3 View FIGURE 3 B). Maxillary apparatus of P-type only, consisting of forceps fused with carrier-like structure and seven pairs of anterior denticles (D). Forceps of adult worms 90–150 µm long, median edge of anterior part of forceps with large robust distal fang and 5–8 large teeth irregularly alternating with small teeth, distal fang much larger than other large teeth ( Fig. 3 View FIGURE 3 C); D1–D3 with distal fang and coarsely serrated median cutting edge; D4–D7 with finely serrated edge, associated with oval ventral plate.
Reproduction and development. Ophryotrocha diadema is hermaphroditic. The first and second chaetigers are sterile, the third and fourth are male, and the remainder are female. The chromosome number is 2n = 8. Newly laid eggs are ovoid, 200 x 180 µm. The egg case is firm and elastic, transparent, and of fusiform shape, closed at both ends. The larvae are released from the egg case at the 4-chaetiger stage.The jaw ontogeny has been described and illustrated on the basis of SEM images by Macnaughton et al. (2010). Larval maxillae are present in up to 5- chaetiger animals, juvenile (P1) maxillae are found only in 6- and 7-chaetiger animals, and adult (P2) maxillae from 7- to 8-chaetiger animals onwards. Other reproductive and population parameters are listed in Table 2 View TABLE 2 and will be compared for the three species in the “Discussion” below.
Remarks. Ophryotrocha diadema was originally collected in August 1972 by Dr. Donald J. Reish (and his daughter) in the harbour of Los Angeles. It proved to be a rare species among those sampled. Only two individuals were brought to Gothenburg together with several hundreds of O. labronica pacifica . In 1989, a second collection of more than 50 specimens was made by Åkesson in a small-boat marina in Long Beach, California. The best site was an old piece of rope, overgrown by mussels and other fouling organisms, with O. diadema living in the interstices of that fouling community. Recently it has been found in the Sicilian harbour of Porto Empedocle ( Simonini et al. 2009, 2010), presumably resulting from anthropogenic activities, e.g., transport of ballast water or in fouling material of ships.
Both the two individuals of 1972 and the population of 1989 carried an egg colour polymorphism, a diallelic mutation with one dominant ( Y) and one recessive allele (w) expressing itself as white or pale yellow. The white eggs of the (ww) homozygotes result from the inability of the worms to synthesize lutein ( Sella & Marzoná 1982). The mutation remains in the population due to a higher fecundity of the heterozygotes ( Åkesson 1976). The YY, Yw, and ww genotypes of O. diadema have been compared in a life table analysis ( Åkesson 1982). In the present comparison performed here we used the wild type data ( Table 2 View TABLE 2 ).
Dr. G. Sella and her team in Torino got an early access to O. diadema with the three genotypes. In one of their major research projects they analysed the ways in which the functional female in a bond can avoid to be cheated by her partner, i.e., that the male deserts the fertilised female and prefers to repeat the male function together with other females ( Sella & Marzoná 1982; Sella 1988; Premoli & Sella 1995; Sella & Lorenzi 2000). Another interesting – and difficult – calculation would be their attempt to determine the sperm redundancy (number of sperm offered per egg). They arrived at the very low figure of 50 spermatozoa per egg ( Sella 1990).
Abiotic factors, such as temperature and salinity and their effect on reproduction, have been studied by Åkesson & Costlow (1978). The optimum salinity is 33‰, the optimum temperature occurs at 25ºC. In order to facilitate comparisons with the two new species, we have run life tables in a combination of 33‰ salinity and 21ºC.
Ophryotrocha diadema View in CoL is one of the best known polychaetes to be used for bioassays. Many details of its life history can be recorded in those assays: (1) time from spawning to larval release; (2) the fate of the embryos inside the transparent egg mass; (3) the time from larval release to spawning of virginal individuals. It is important, however, to know the genetic properties of the strain used for assays ( Åkesson 1982). Reish & Carr (1978) used O. diadema View in CoL in an analysis of heavy metals. Hooftman & Vink (1980) reported on toxic effects of dieldrin and two other chemicals. They compared short-term and long-term effects. Klöckner (1979) studied effects of cadmium on three consecutive generations. Recently, Knowles & Greenwood (1994) used seven continuous generations of O. diadema View in CoL to study effects of doses of gamma radiation.
Distribution. Pacific Ocean: Los Angeles and Long Beach, California; Mediterranean Sea: Porto Empedocle, Sicily.
O. diadema O. alborana , sp.nov. O. birgittae , sp. nov.


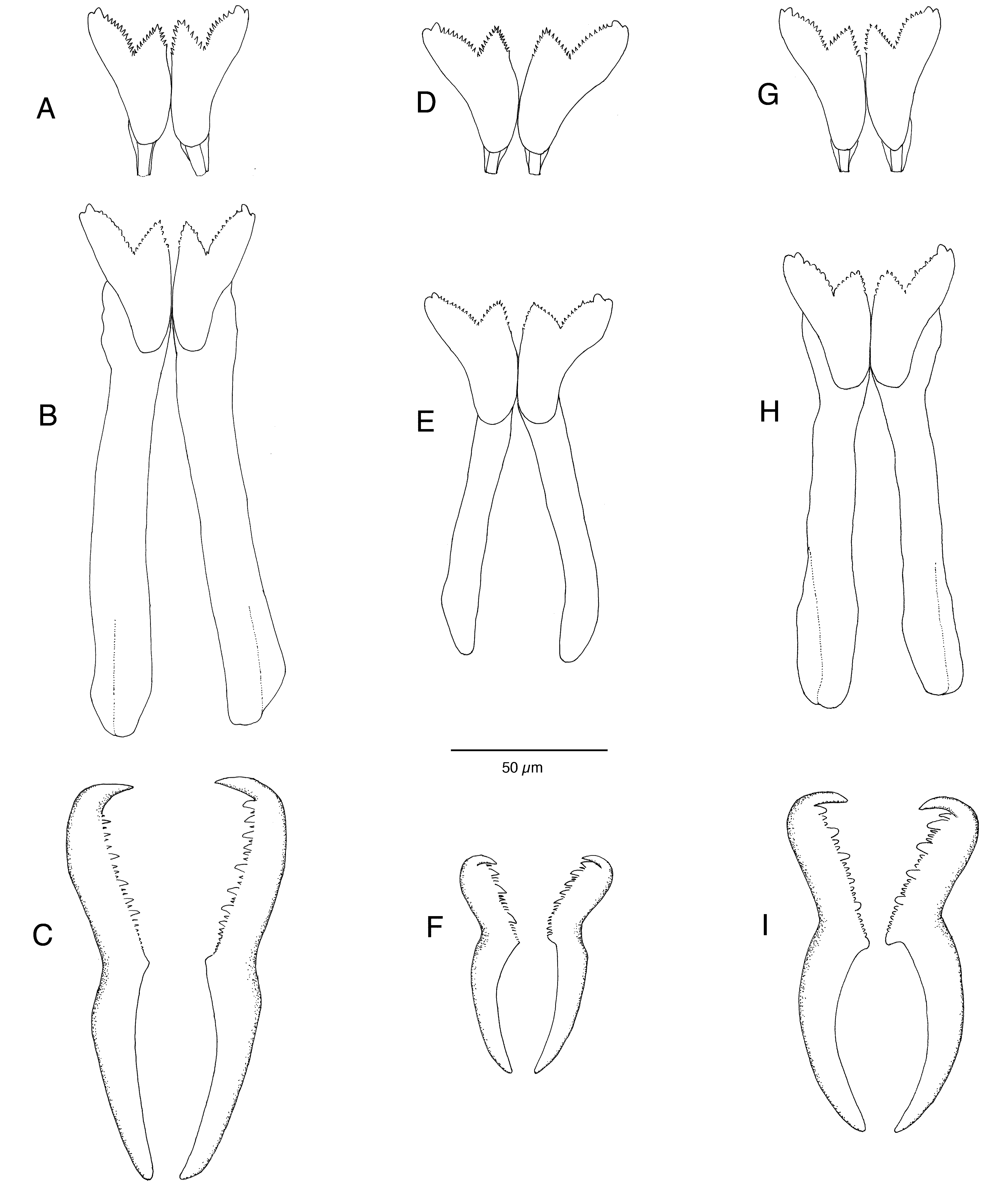
TABLE 1. Comparison between species of the Ophryotrocha diadema group.
| Feature | O. diadema | O. alborana , sp.nov. | O. birgittae , sp. nov. |
|---|---|---|---|
| Morphology | |||
| Length of most live adults | 2.5–3.5 mm (15–17 chaetigers) | 3.0–4.0 (17–20 chaetigers) | 3.5–4.5 (17–19 chaetigers) |
| Maximum length of live adults | 4.6 mm (25 chaetigers) | 5.0 (25 chaetigers) | 5.5 (23 chaetigers) |
| Eyes | Two connected eyes appear- ing as silvery white triangle | Two separate silvery white eyes with additional red pig- ment cup | Two separate, silvery white eyes |
| Rosette glands on posterior seg- ments | Up to 5 pairs | Up to 9 pairs | Up to 9 pairs |
| Shape of mandibles of adult worms | Shafts with basal and distal lateral thickenings | Slender shafts | Shafts with basal and distal lateral thickenings |
| Anterior edge of mandibles | With 22–28 tiny teeth/plate | With 22–28 tiny teeth/plate | With 20–24 tiny teeth/plate |
| Length of mandibles of adult worm | 110–170 µm | 100–145 µm | 125–140 µm |
| Distal fang of maxillary forceps | Robust and much larger than other large teeth | Slightly larger than other large teeth | Slender but much larger than other large teeth |
| No. of large teeth on maxillary forceps | 5–8 | 3–5 | 3–5 |
| Length of forceps of adult worm | 90–150 µm | 60–85 µm | 90–125 µm |
| Reproduction and development | |||
| Size of eggs | 200 x 180 µm | 190 x 150 µm | 200 x 160 µm |
| No. of chaetigers at larval release | 4 | 4–5 | 4 |
| Presence of larval maxillae | 4 to 5 chaetiger worms | 5 to 6 chaetiger worms | 4 to 6 chaetiger worms |
| Presence of juvenile maxillae (P1) | 6 to 7 chaetiger worms | 7 to 11 chaetiger worms | 7 to 11 chaetiger worms |
| Presence of adult maxillae (P2) | 7 to 8 chaetiger worms onwards | 8 to 12 chaetiger worms onwards | 12 chaetiger worms onwards |
| SMNH |
Saskatchewan Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ophryotrocha diadema Åkesson, 1976
| Paxton, Hannelore & Åkesson, Bertil 2011 |
Ophryotrocha diadema Åkesson, 1976 : 23
| Akesson 1976: 23 |