
Bathyceramaster elegans ( Ludwig, 1905 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4175.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:B264C215-000D-42C5-8AC9-B801872CD182 |
|
DOI |
https://doi.org/10.5281/zenodo.6055201 |
|
persistent identifier |
https://treatment.plazi.org/id/953787B9-FF88-FFE3-5CF0-F8938052F904 |
|
treatment provided by |
Plazi |
|
scientific name |
Bathyceramaster elegans ( Ludwig, 1905 ) |
| status |
|
Bathyceramaster elegans ( Ludwig, 1905) View in CoL
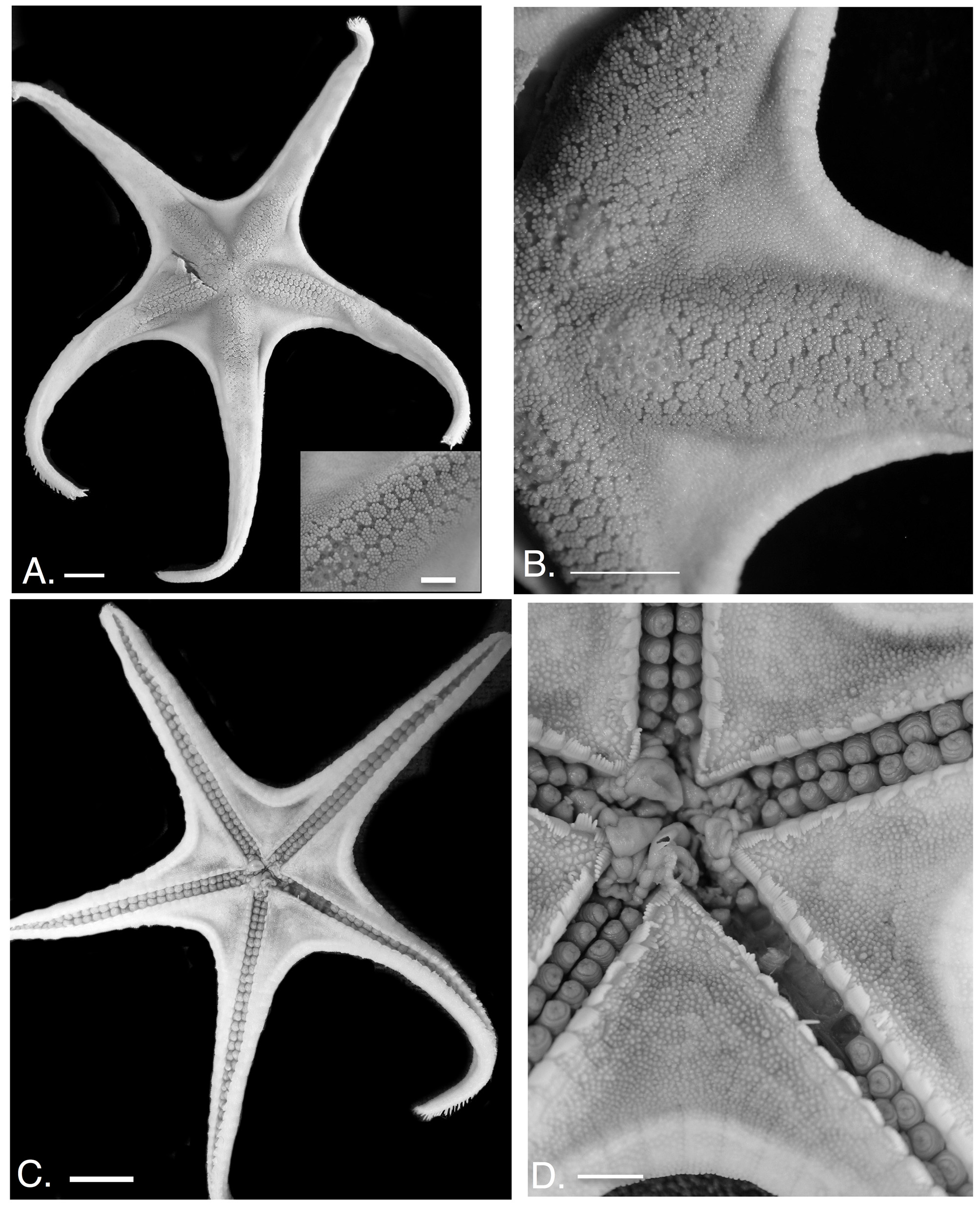
Figures 2 View FIGURE 2 A–G (holotype), 3A–D (new material)
Mediaster elegans Ludwig 1905: 125 View in CoL ; Fisher, 1911: 197, 204; Madsen 1961b: 88. Mediaster elegans abyssi Ludwig 1905: 128 .
Comments. The species presented as “ Mediaster elegans ” from Carey (1972) and Maluf (1988) is described herein as Bathyceramaster careyi . Material outlined from those citations was compared with Ludwig’s (1905) type and discovered to be incorrectly identified. Thus, “ Mediaster ” elegans individuals consistent with the holotype ( Fig. 2 View FIGURE 2 A–G) has not been observed since its description by Ludwig (1905). This represents a new and novel range occurrence for this species.
Newly collected material of Bathyceramaster elegans showed some variation relative to Ludwig’s holotype. Granulation and abactinal plate patterns differed between the holotype, which is the largest specimen of this species known, and the more recently collected newer individuals. Also different was the presence of pedicellariae in the holotype and the two specimens collected from Baja California but absent from more northern individuals.
One other significant difference was that the disk and arms of the holotype show the abactinal surface of the body as much more inflated than any of the newly collected specimens, which all have relatively flattened body surfaces. However, upon comparing the holotype with individuals observed in situ, it became clear that when alive, this species displays a very swollen abactinal disk surface filled with seawater. Thus, it seems that this character difference is best explained as a collection artefact dependent on whether the body coelom swells with water or is evacuated during the preservation process.
Ludwig (1905) described Mediaster elegans abyssi as a variety of M. elegans based on its fewer number of furrow spines (six or seven versus nine or ten in the Cocos Island holotype).
Based on ICZN regulation 45.6.4, based on this term’s usage as “var.”, prior to 1961, the epithet abyssi is automatically considered a subspecies (as used by Carey 1972). It seems likely that the furrow spine number is likely associated with this specimen’s smaller size (R=4.5 versus 8.2 in the holotype). Although, Ludwig’s type for this subspecies could not be located in either in the NMNH collections (e.g., Ahearn 1995) or in the collections of the California Academy of Sciences (e.g., Chaffee & Weitbrecht 1984), his description does not allude to any significant differences and thus seems most likely a synonym of M. elegans .
A re-description of this species is presented herein, incorporating information from the holotype, the new specimens collected and in situ observations.
Occurrence. Co-Axial Cone, President Jackson Seamount B and C, North Pacific Ocean to Alarcon Rise, Baja California to Cocos Island, Costa Rica, Bay of Panama. 1790–3335 m.
In situ observations. North Pacific specimens with strongly swollen disks, presumably filled with seawater, were observed on basalt substrates covered with sediment.
Description. Disk thick, body strongly stellate (R/r=3.7–4.9) with arms, long tapering; triangular to elongate in shape. Interradial arcs broadly curved to straight. Holotype with swollen disk and arms ( Fig. 2 View FIGURE 2 A). Body relatively flattened in newly collected specimens ( Fig. 3 View FIGURE 3 A).
Abactinal plates polygonal in outline ( Fig. 2 View FIGURE 2 A,B; 3A,B), ranging from wide (W>L) to equal (L=W) in diameter. Plates are larger on disk and on basal arm region becoming smaller (> 50%) distally especially near armtips and adjacent to superomarginals. Bases of abactinal plates polygonal to quadrate ( Fig. 2 View FIGURE 2 C). Radial plate series, especially the carinal series differentiated from those interradially. Radial plates tend to be wider (L>W) in most individuals examined ( Fig. 3 View FIGURE 3 A inset). The holotype (R=8.2) shows multiple rows of wide plates (L>W) on the disk/basal arm regions with the plate dimensions narrowing distally ( Fig. 2 View FIGURE 2 A, B). A smaller individual (R= 5.3) showed poorer differentiation between plates along these different regions and displays more round to polygonal plates between radial and interracial regions. Larger individuals show greater propensity for ordered, continuous fasciolar channels between radial and interradial regions whereas those in smaller individuals appear to display less continuity. Granules, 10–60, mostly 20–30, coarse, polygonal to quadrate in outline. Most disk granules ( Fig. 2 View FIGURE 2 A inset, B; 3A inset, B) differentiated onto central and peripheral areas. Central granules round or polygonal, variable one to 20, mostly four to 18. Peripheral granules mostly quadrate/rectangular, variably four to 40, mostly ten to 30, similar or up to twice the size of central granules ( Figs. 2 View FIGURE 2 A,B; 3A,B). Granule distribution ranges from densely obscuring plate boundaries to widely spaced evenly on a plate. Interradial regions with more crowded granules in most individuals. Larger individuals with more crowded, densely arranged granules versus smaller individuals which are more widely and evenly spaced apart ( Fig. 2 View FIGURE 2 A vs.3A).
Pedicellariae clam shell like, bivalve or trivalve, each about 0.5 mm or about the size of two adjacent granules. Madreporite shape ranges from triangular, quadrate to polygonal and round flanked by three to five adjacent plates. Papulae distinct, approximately six per plate on radial/proximal regions, absent interradially. No pedicellariae observed on abactinal surface in the North Pacific specimens, but simple clam-shell type pedicellariae, similar in Marginal plates facing laterally, approximately 72–76 per interradius (armtip to armtip). Superomarginals quadrate in outline interradially (L=W), becoming more elongate and slightly more irregular in shape distally (L>W). Inferomarginals wider interradially becoming more elongate distally. Marginal plate surface covered by round to polygonal granules, 70–200, densely arranged identical to those on abactinal surface and continuous with disk covering ( Fig. 2 View FIGURE 2 D, 3B). Granules form discrete periphery on each marginal plate series, approximately 15x15 on superomarginals. Inferomarginals with length equal to approximately 15 granules and width equal to approximately 20 granules. Superomarginals covered only by granules. Individuals collected from the North Pacific all with a large, nubbin-like or sharp-tipped spine present on central surface of inferomarginal plates. This spine is absent from the more southern holotype. Many of the inferomarginals on the holotype are denuded, making it unclear if a spine or spines were present. Pedicellariae on marginals absent from North Pacific specimens, but present on those from Baja California. When present, pedicellariae number one or two, identical to those on abactinal surface present on nearly every marginal plate. Terminal plate small, triangular in shape, smooth surface.
Actinal regions large, plates quadrate/polygonal to irregular in shape, arranged in two to three irregular chevron like series ( Fig. 2 View FIGURE 2 E, F). Plate surface covered by angular to polygonal granules, five to 30 variably distributed. Larger individuals with granules/spinelets more associated with discrete plates, whereas smaller, North Pacific specimens with evenly distributed granules/spines across the actinal surface regardless of plate boundary. When discrete plates are present, peripheral granules, eight to fifteen more elongate and angular in shape. Shallow fasciolar grooves present in larger individuals. Central granules one to eight. Granules widely and evenly spaced proximally becoming more crowded and densely arranged distally adjacent to the inferomarginals. Pedicellariae identical to those on abactinal and superomarginal surfaces also present on actinal plate surfaces but mostly adjacent to inferomarginals. Proximal, actinal region with few to no pedicellariae. Where pedicellariae have been removed, a shallow pit (approximately 1–3 mm deep) remains.
Furrow spines 9 to 15, mostly 11, with number increasing only near armtip. Spines slender, blunt tipped, arranged in a weakly curved fan ( Fig. 2 View FIGURE 2 G, 3C, D). Subambulacrals three to six, each approximately three to five times as thick as the furrow spines with largest in center of series, shortest, smallest at end of series. Pedicellariae behind subambulacral spines/granules, large, more elongate than furrow spines, valves at least twice as thick as each furrow spine. Where pedicellariae are lost, large depressions present. Subsequent spines/granules on adambulacral plate surface identical to those elsewhere on actinal surface. Furrow spines, all similar in height, blunt tipped, on oral plates, 16–20 on each side meeting at sharp angle projecting into the mouth. Oral plate surface with nine paired angular granules/spinelets on each half of the oral plate ( Fig. 2 View FIGURE 2 E,F,G; 3D). Approximately three to six remaining angular granules/spinelets on each oral plate surface. Pedicellariae absent in North Pacific individuals, but present in those from Baja California. When present pedicellariae are bivalve or trivalve and are similar in size to two or three granules (about 0.5 mm if measured along a line). Pedicellariae present on nearly every actinal plate in Baja specimens. Tube foot furrow wide and distinct with muscular tube feet.
Color in life is white to light yellow/orange.
Material examined. USNM 34443 About USNM (Holotype), Cocos Island , Costa Rica, 5º43”N 85º50”W, 1789 m, Coll GoogleMaps . USFC Steamer Albatross 1 wet spec. R=8.2 r=2.2; North Pacific . USNM 1407937 About USNM , Co-Axial Cone, North Pacific. 45.517, -129.583, 2406.5 m, Coll. D. Clague et al. , MBARI. St. D77-A2, 1 wet spec. R=6.3, r=1.6, USNM 1407938 About USNM , President Jackson Seamount B, North Pacific. 42.83, -128.159, 2019.3m, Coll. D. Clague et al . MBARI, St. D81-A6. 1 wet spec.R=7.1, r=1.8. USNM 1407939 About USNM , President Jackson Seamount C, North Pacific. 42.7439, - 128.0984, 1801.3 m, Coll. D. Clague et al . MBARI, St. D 82-A2. 1 wet spec. R=6.4, r=1.3. Baja & South America . USNM 1407940 About USNM , Alarcon Rise , Baja California, 22.377036, -108.531294, 2289.8 m. Coll. D. Clague et al. , MBARI, D398-A4.1 wet spec. R=5.3, r=1.3, (arms upturned). USNM 1407941 About USNM , Alarcon Rise , Baja California, 23.377024, -108.531334, 2290.1 m. Coll. D. Clague et al. , MBARI. D398-A2, 1 wet spec. R=7.1, r=1.8.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bathyceramaster elegans ( Ludwig, 1905 )
| Mah, Christopher L. 2016 |
Mediaster elegans
| Madsen 1961: 88 |
| Fisher 1911: 197 |
| Ludwig 1905: 125 |
| Ludwig 1905: 128 |