
Anolis nebulosus ( Wiegmann 1834 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3862.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3FA375FE-E4E0-4509-BE02-EE5E786B07C6 |
|
DOI |
https://doi.org/10.5281/zenodo.7534614 |
|
persistent identifier |
https://treatment.plazi.org/id/8A288798-FFB4-E746-7EC3-FA30FC17C19C |
|
treatment provided by |
Felipe |
|
scientific name |
Anolis nebulosus ( Wiegmann 1834 ) |
| status |
|
Anolis nebulosus ( Wiegmann 1834)
Figs. 95–101 View FIGURE 95 View FIGURE 96 View FIGURE 97 View FIGURE 98 View FIGURE 99 View FIGURE 100 View FIGURE 101
Dactyloa nebulosa Wiegmann 1834: 47 View in CoL ; type locality: “ Mexico ”; restricted to Mazatlán, Sinaloa by Smith & Taylor 1950b. Holotype: ZMB 527.
Anolis nebuloides: Oliver 1937 , Bogert & Oliver 1945, Zweifel & Norris 1955, Smith & Grant 1958, Tanner & Robinson 1959, Fugler & Dixon 1961
Anolis nebulosis: Fosdick 1968
Anolis nebulosus: Lichtenstein 1856 , Jan 1857, Cope 1866, Dugès 1869, Bocourt 1873, O’Shaughnessy 1875, Dugès 1878, Cope 1879, Müller 1884, Günther 1885 –1902, Cope 1886, Cope 1887 (in part.), Dugès 1889, Cope 1896, Dugès 1896, Van Denburgh 1897, Mocquard 1899, Stejneger 1899, Cope 1900, Herrera 1904, Boulenger 1905, Gadow 1905, 1910, Van Denburgh & Slevin 1914, Slevin 1926, Barbour 1934 (in part.), Taylor 1936, Smith 1939, Burt & Myers 1942, Martín del Campo y Sánchez 1942, Smith 1943, Smith & Mittleman 1943, Schmidt & Shannon 1947, Stuart 1948, Smith & Taylor 1950a, b, Davis & Smith 1953, Davis 1954, Duellman 1954, Peters 1954, Lewis & Johnson 1955, Duellman 1956, Lewis & Johnson 1956, Gehlbach & Colette 1957, Drake 1958, Duellman 1958a, b, Malkin 1958, Smith & Grant 1958, Duellman & Duellman 1959, Etheridge 1959, Zweifel 1959, Grant & Smith 1960, Peters 1960, Zweifel 1960, Davis & Dixon 1961, Duellman 1961, Webb & Baker 1962, Anderson & Lidicker 1963, McDiarmid 1963, Etheridge 1965, Duellman 1965, Dixon & Webb 1966, Lynch & Smith 1966, Jenssen 1968, Bellaris 1969, Conant 1969, Hardy & McDiarmid 1969, Jenssen 1969, Smith 1969, Taylor 1969, Webb & Baker 1969, Fitch 1970, Jenssen 1970a,b, 1971, Murphy 1971, Purdue & Carpenter 1972, Greene 1972, Smith et al. 1972, Crossin et al. 1973, Smith & Smith 1973, Fitch & Henderson 1973, Fitch 1976, Smith & Smith 1976, Lieb 1981, Fitch & Hillis 1984, Gorman et al. 1984, Tanner 1987, Flores-Villela & Hernandez-García 1989, McCranie & Wilson 1990, Lister & García Aguayo 1992, Flores-Villela 1993, Flores-Villela & Gerez 1994, Lieb 1995, Mayén-Peña & Salgado-Maldonado 1998, Pérez-Ramos et al. 2000, Lieb 2001, Ramírez-Bautista & Benabib 2001, Bryson et al. 2002, Gienger et al. 2002, Ramírez-Bautista & Pardo-De La Rose 2002, Poe 2004, FloresVillela & Hernandez-García 2006, Santamaría & Flores-Villela 2006, Boyd et al. 2007, Flores-Villela & CansecoMárquez 2007, Liner 2007, García 2008, Fläschendräger & Wijffels 2009, Lemos-Espinal & Smith 2009, Losos 2009, Sanchez-Luna et al. 2009, Castigilia et al. 2010, Flores-Villela et al. 2010, Lavín-Murcio & Lazcano 2010, UrbinaCardona & Flores-Villela 2010, Wilson & Townsend 2010, Nieto Montes de Oca et al. 2013, Siliceo-Carentero & García 2013, Wilson et al. 2013, Köhler 2014, Köhler et al. 2014
Anolis schmidti Smith 1939: 21 ; type locality: “Manzanillo, Colima,” Mexico. Holotype: FMNH 1667. Smith & Taylor 1950 a,b, Davis 1954, Duellman 1954, Peters 1954, Stuart 1955, Marx 1958, Etheridge 1959, Peters 1960, Duellman 1961, 1965, Etheridge 1965, Fitch & Henderson 1973, Smith & Smith 1976, Flores-Villela 1993, Flores-Villela & Gerez 1994, Lieb 1995, Liner 2000, Lieb 2001, Santamaría & Flores-Villela 2006, Liner 2007, Urbina-Cardona & Flores-Villela 2010, Wilson & Townsend 2010, Wilson et al. 2013, Nieto Montes de Oca et al. 2013
Anolius nebulosus: Herrera 1899 , Smith & Smith 1975
Norops nebulosus: Savage & Guyer 1989 , McCranie & Wilson 2001, Nicholson 2002, Castigilia et al. 2010, Nicholson et al. 2012
Norops schmidti: Savage & Guyer 1989 , Liner 2000, Nicholson 2002, Nicholson et al. 2012
Diagnosis. A small species (SVL in largest male 43.0 mm, largest female 39.5 mm) of the genus Anolis (sensu Poe 2004) that differs from all Mexican and Central American anoles except A. carlliebi and A. sacamecatensis by having a combination of (1) weakly to strongly keeled ventral scales; (2) usually a patch of three greatly enlarged supraocular scales; (3) 8–18 rows of slightly enlarged dorsal scales that are smaller than ventral scales; (4) short hind legs, longest toe of adpressed hind leg usually reaching to ear opening, exceptionally to a point between ear opening and eye, ratio shank length/SVL 0.21–0.25; (5) a single elongate prenasal scale; and (6) male dewlap in life orange with or without a white anterior margin. Among the Anolis species treated in this contribution, A. nebulosus is most similar to A. microlepidotus from which it differs in male dewlap coloration (uniform orange or orange with a white anterior margin in A. nebulosus vs. dull orange yellow with paler semicircular streaks and blotches in A. microlepidotus ) and larger dorsal scales (number of dorsal scales in one head length 26-38, mean 30.0, in A. nebulosus vs. 32–48, mean 41.2, in A. microlepidotus ).
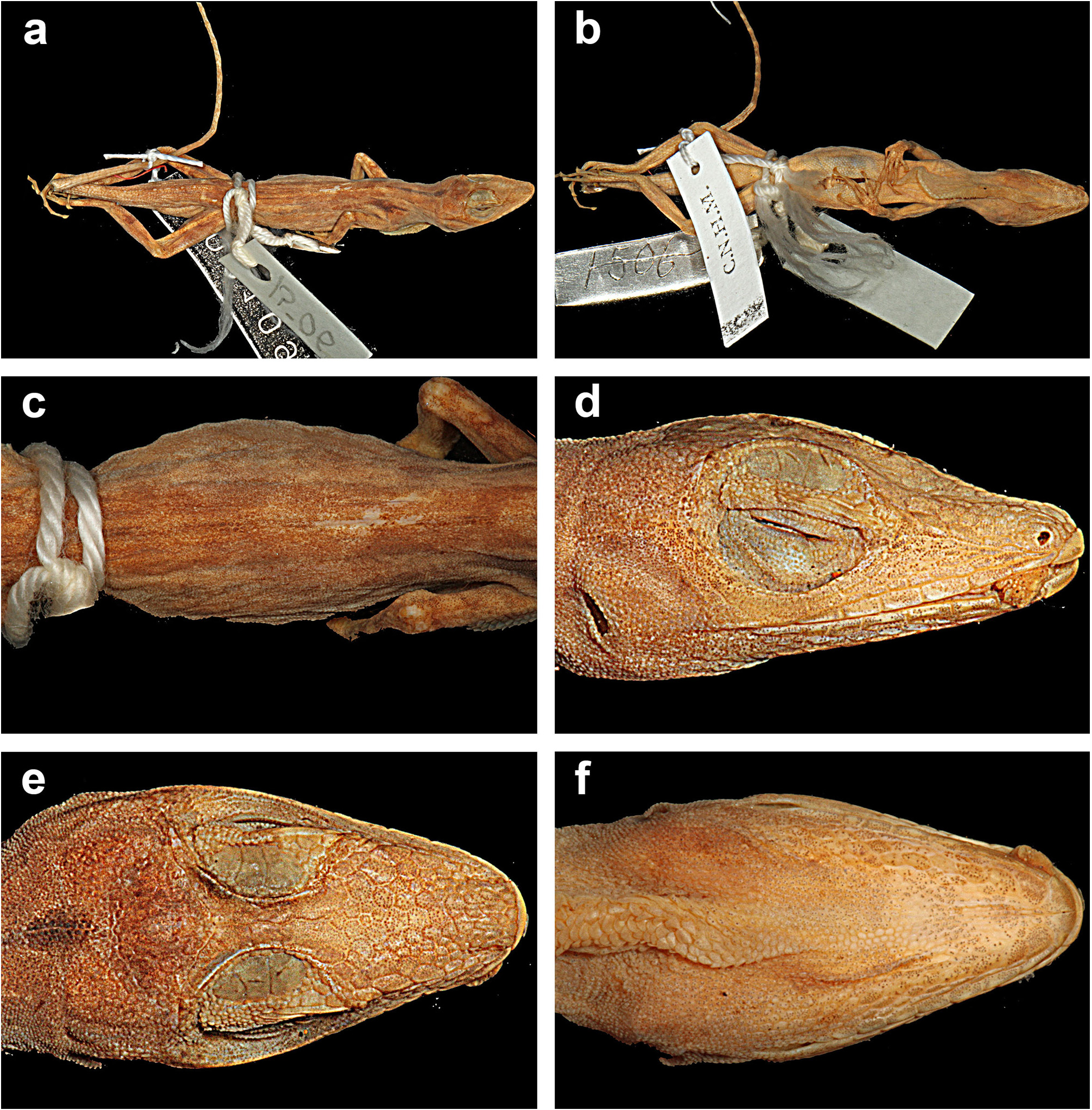
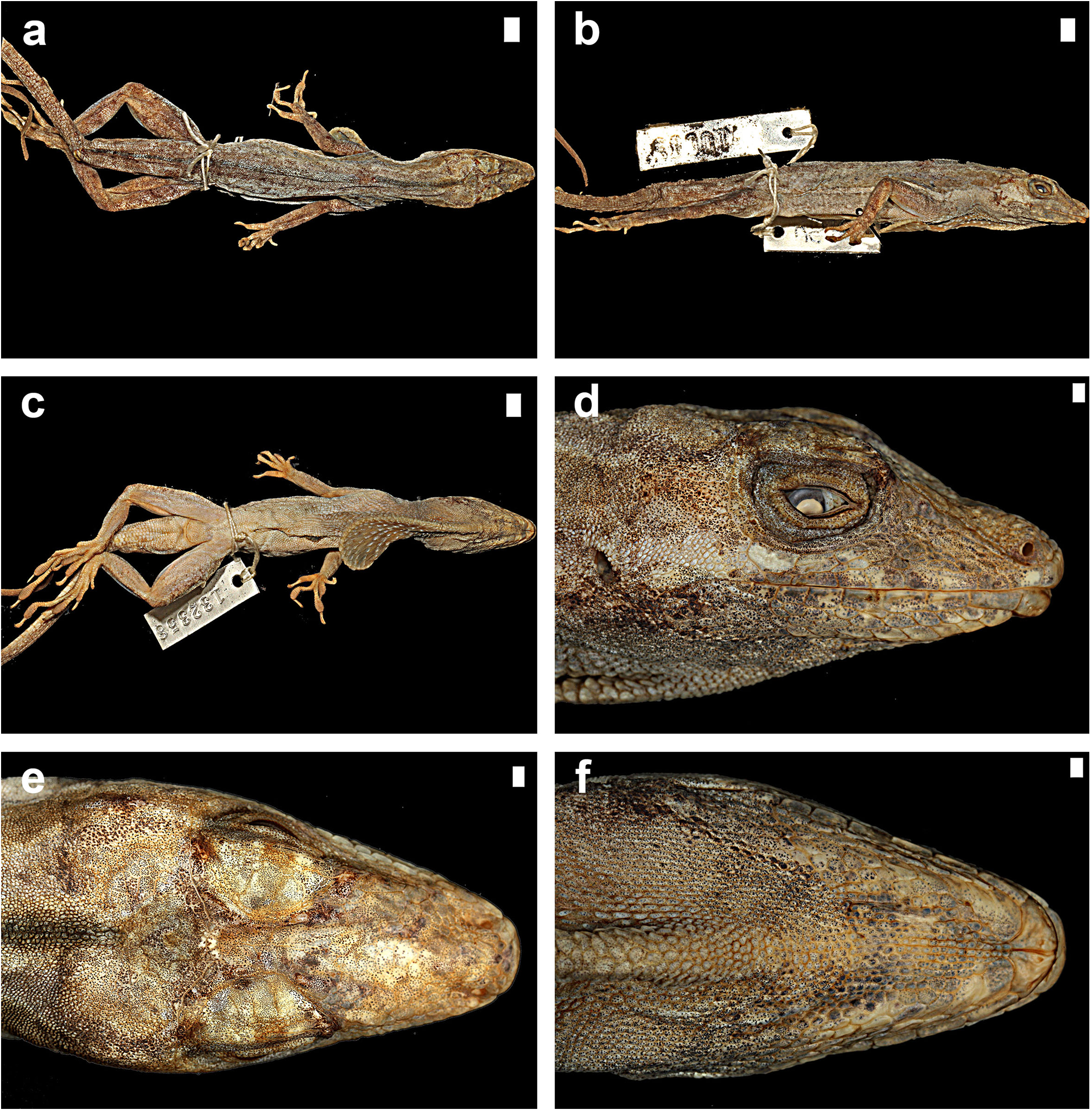
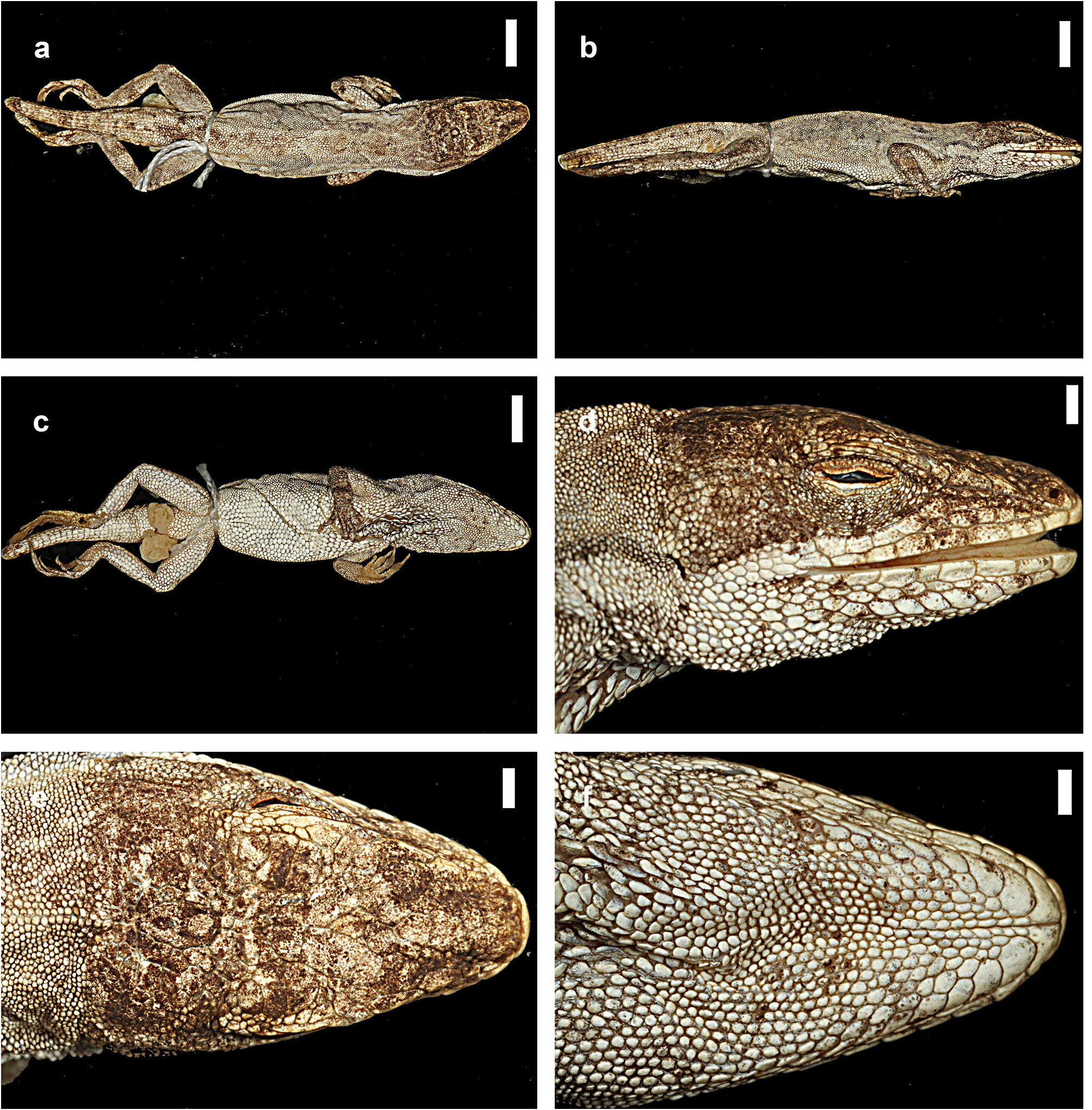
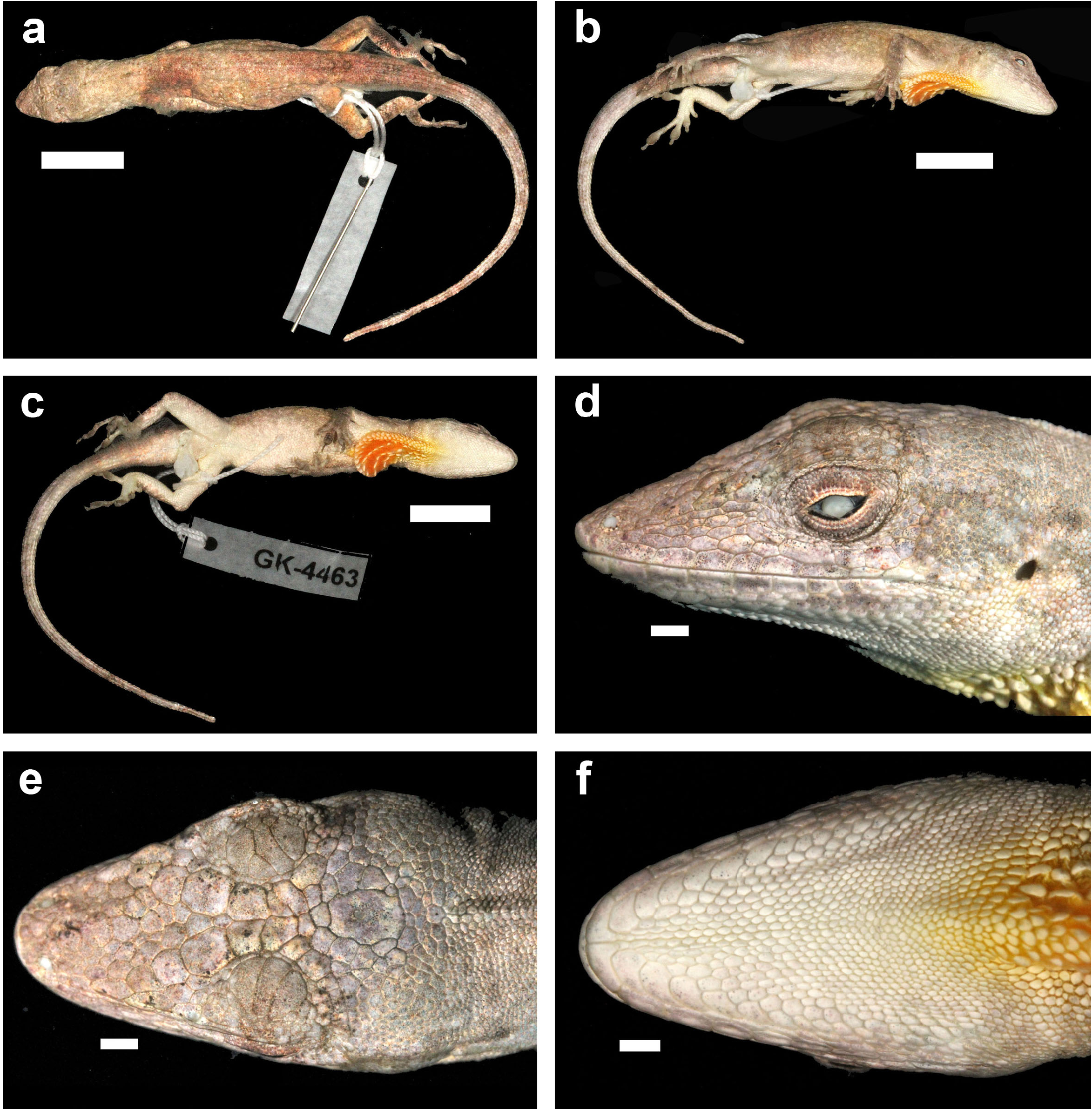
Description. Anolis nebulosus is a small anole (maximum recorded SVL 43.0 mm in males, 39.5 mm in females); dorsal head scales in internasal region mostly or keeled, in prefrontal, parietal, and frontal areas smooth or rugose; moderately deep prefrontal depression present, shallow parietal depression; 4–6 postrostrals; anterior nasal usually single, occasionally divided, the lower scale in contact with rostral and first supralabial; 6–7 internasals; canthal ridge sharply defined; scales comprising supraorbital semicircles well defined, posterior ones smooth, anterior ones with a rounded keel, largest scale in semicircles about subequal to largest supraocular scale; supraorbital semicircles usually broadly in contact, occasionally separated by one scale row; 1–3 scales separating supraorbital semicircles and interparietal at narrowest point; interparietal well defined, greatly enlarged relative to adjacent scales, surrounded by scales of moderate size, usually longer than wide, much larger than ear opening; enlarged supraoculars usually smooth, occasionally rugose, usually broadly in contact exceptionally separated from supraorbital semicircles by a complete row of small scales; 1–2 scales between enlarged supraoculars and superciliaries; 2 elongate superciliaries, anterior one longest, followed posteriorly by a series of 3–4 moderatesized keeled, rounded or squarish scales; usually 3 enlarged canthals, the second canthal largest; 4–6 scales between second canthals; 6–10 scales present between posterior canthals; loreal region slightly concave, 14–26 mostly keeled (some smooth or rugose) loreal scales in a maximum of 4–5 horizontal rows; 6–8 supralabials to level below center of eye; suboculars keeled, in broad contact with supralabials (3–4 suboculars in contact with 2–5 supralabials); ear opening vertically oval, oriented slightly obliquely; scales anterior to ear opening granulars, about twice as large as those posterior to ear opening; 6–9 infralabials to level below center of eye; 4–5 (usually 4) postmentals, outer pair usually at least four times larger than adjacent median postmental scales; 0–2 (usually 1, commonly 2) enlarged sublabials in contact with infralabials on each side; weakly to strongly keeled granular scales present on chin and throat; male dewlap of large size extending from level below anterior margin of eye to level of chest; 8–9 horizontal gorgetal-sternal rows with 8–14 scales per row; modal number of marginal pairs 2–4; female dewlap very small; a nuchal crest and a dorsal ridge present in males; scales on middorsum weakly to moderately keeled, subimbricate; scales on middorsum keeled, juxtaposed to subimbricate with rounded posterior margins; 8–18 middorsal scale rows slightly enlarged, with a gradual transition to lateral scales; lateral scales keeled, granular, usually somewhat heterogeneous, with some slightly enlarged conical scales scattered among granulars; 50–62 dorsal scales along vertebral midline between levels of axilla and groin in males, 55–71 in females; 26–38 dorsal scales along vertebral midline contained in one head length in males, 26–30 in females; ventral scales on midsection about twice the size of largest dorsal scales; scales on midventer distinctly keeled, juxtaposed to subimbricate, usually with rounded posterior margins, occasionally slightly mucronate; 46–62 ventral scales along midventral line between levels of axilla and groin in males, 52–54 in females; 32–40 ventral scales contained in one head length in males, 26–30 in females; 108–136 scales around midbody in males, 110–124 in females; tube-like axillary pocket absent; precloacal scales weakly keeled; males with a pair of moderately to greatly enlarged postcloacal scales; tail moderately compressed in cross section, tail height/tail width 1.11–1.50 in males, 1.13–1.42 in females; basal caudal scales faintly to weakly keeled, all other caudal scales strongly keeled; lateral caudal scales, homogeneous although an indistinct division in segments is discernible; dorsal medial caudal scale row not enlarged; scales on anterodorsal surface of brachium subimbricate to imbricate, strongly keeled, unicarinate, some slightly mucronate; scales on dorsal surface of antebrachium strongly keeled, unicarinate, subimbricate to imbricate, non-mucronate; 19–23 subdigital lamellae on Phalanges II–IV of Toe IV of hind limbs; 4–6 subdigital lamellae on distal phalanx of Toe IV of hind limbs; digital pads dilated, slightly more than twice the size of distal phalanx. In all specimens examined, the longest toe of the adpressed hind leg reaches to level of tympanum or to a point between shoulder and tympanum. For variation in selected scalation and morphometric characters see Table 11 View TABLE 11 .
The coloration in life of an adult male from Acapulco (SMF 96221) was recorded as follows: Dorsal ground color Drab (19) with indistinct Dark Drab (45) chevrons and an Army Brown (46) interorbital bar; lateral surface of body Clay Color (18); Dorsal surface of tail Drab (19) with indistinct Clay Color (18) bands; dorsal surface of hind limbs Olive Brown (278) with Burnt Umber (48) bands; chin Cream White (52) with Grayish Horn Color (268) mottling; ventral surface of body, limbs and tail Chamois (84) but paler with Grayish Horn Color (268) stipples: dewlap ground color Orange Yellow (8) fading into a small Tawny (60) blotch in center and with a Cream White (52) semicircle bordering the frontal edge of the dewlap; iris Raw Umber (280).
The coloration in life of an adult male from Agua de Obispo (SMF 96223) was recorded as follows: Dewlap ground color Light Orange Yellow (77) with Pratt´s Rufous (72) oblique lines between gorgetals; gorgetals Cream White (52); iris Brussels Brown (33).
The almost completely everted hemipenis of SMF 96221 ( Fig. 101 View FIGURE 101 ) is a small organ with rudimentary lobes; sulcus spermaticus bordered by well developed sulcal lips and opening into a single small apical field void of ornamentation; a finger-like processus present near tip of apex; no surface ornamentation discernable.
Natural History Notes: In the area of Agua de Obispo, we collected Anolis nebulosus mostly during our night searches where we encountered these lizards sleeping on low vegetation along the trails at 0.3 to 1.0 m above the ground. In the archaeological site Palma Sola at Acapulco, we observed one specimen crossing the trail in the afternoon. Anolis nebulosus has been shown to vary its foraging sites and feeding rate with seasonal variations in prey abundance ( Lister and García Aguayo 1992). Also, preferred niches can vary in the course of the day as demonstrated by the observations by García (2008): The use of litter by Anolis nebulosus was relatively higher during the first hours of the day, whereas this species occupied both low and high perches in similar proportions during the rest of the day. Jenssen (1970a) reported the mean body temperature in A. nebulosus to be 29.7 °C. The lizards basked in the early morning hours (06.00–10.00h) and then retired into the shade. The anoles were least active during the warmest part of the day (11.00–15.00h), at which time their body temperatures were over 31.0 °C. Boyd et al. (2007) reported observations on the stomach contents of 8 specimens of A. nebulosus from Chínipas, Chihuahua, and found that numerically, termites were the dominant prey, but volumetrically grasshoppers were most important. The reproductive season mainly coincides with the wet season ( Jenssen 1970a). Field observations suggest a population turnover of about one year, i.e., few individuals appeared to live more than a year in nature ( Jenssen 1970a).
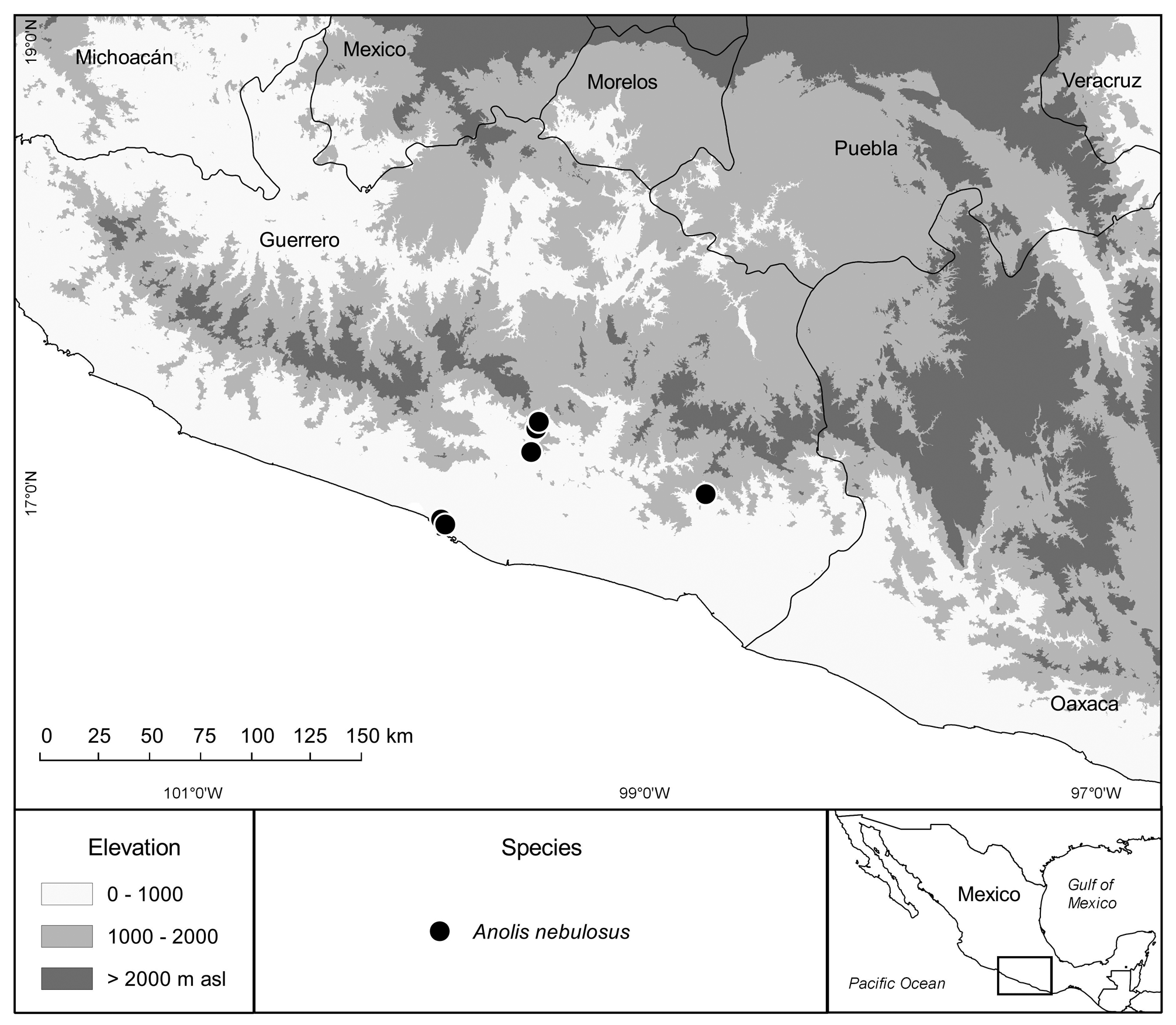
Geographic Distribution and Conservation. As currently known, Anolis nebulosus is distributed along the Pacific versant from the western portion of the Mexican State of Oaxaca to the northern portions of the Mexican States of Sonora and Chihuahua, respectively; in the state of Guerrero it occurs at elevations between 10 and 960 masl ( Fig. 102 View FIGURE 102 ). The taxonomy of this species complex needs to be clarified before the conservation status can be assessed.
Specimens examined ⎯ Mexico: “ Mexico ”: ZMB 527 View Materials ; Guerrero: Acapulco : USNM 46680 About USNM ; Acapulco, Zona Arqueológica Palma Sola , 330 m: SMF 96221 ; Agua de Obispo , 960 m: FMNH 122008 About FMNH , IBH 26551–53 View Materials , 26558 View Materials , SMF 96223–25 About SMF , 96433 About SMF , UIMNH 20135 View Materials , 20137–38 View Materials ; Cascada de Santa Cruz El Rincón , 622 m: SMF 96434–37 About SMF ; El Rincón : UIMNH 20133 View Materials ; Sitio Arqueológico Tehuacalco , 635 m: SMF 96222 .
Taxonomy of the Mexican anoles related to Anolis liogaster Boulenger 1905
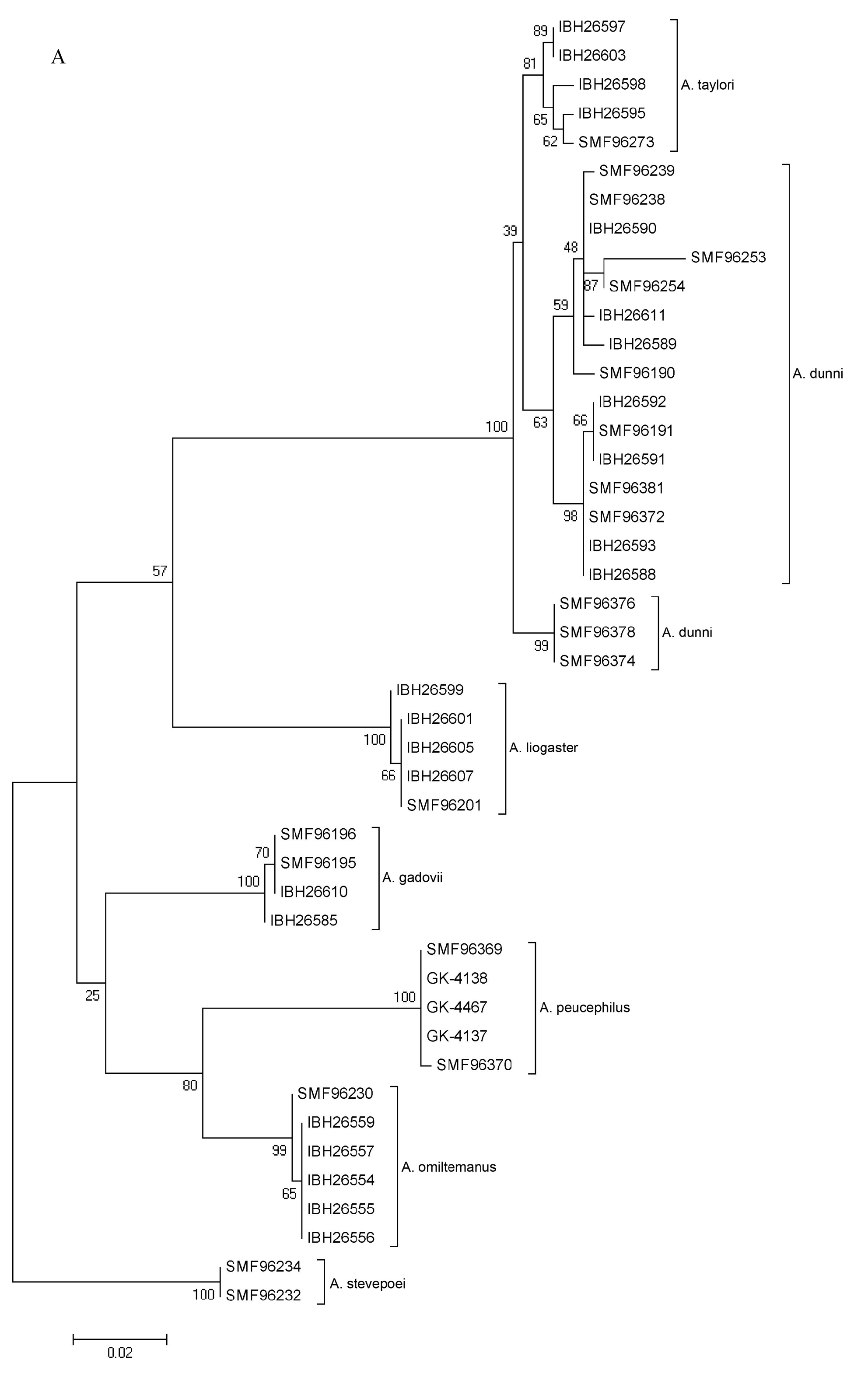
The species of anoles related to Anolis liogaster (i.e., A. dunni , A. gadovii , A. liogaster , A. omiltemanus , A. peucephilus , and A. taylori ) are readily differentiated from all other Mexican and Central American congeners by having (1) smooth ventral scales; (2) an oval patch of usually three greatly enlarged supraorbital scales; (3) a pair of greatly enlarged postcloacal scales in males. In 1905, Boulenger described two new species of anoles: Anolis liogaster based on a series of specimens (syntypes: BMNH 1946.8.8.53–54; Fig. 103 View FIGURE 103 ) from “Omilteme, Guerrero, 7600 ft. ” and Anolis gadovii based on an adult male specimen (holotype: BMNH 1946.8.13.1; Fig. 104 View FIGURE 104 ) from “Tierra Colorada, South Guerrero.” Smith (1936) described Anolis dunni based on a specimen (now FMNH 100109; Fig. 105 View FIGURE 105 ) from “on a boulder in the high mountains, within the evergreen zone, between Rincón and Cajones, Guerrero.” Smith & Spieler (1945) described Anolis taylori from “the hills about one mile north of Acapulco, Guerrero ” (holotype: USNM 132358; Fig. 106 View FIGURE 106 ). In 1954, Davis described Anolis omiltemanus (holotype: TCWC 10278; Fig. 107 View FIGURE 107 ) from “two miles west of Omiltemi, 7800 ft., Guerrero.” Smith (1972) named his new species, Anolis adleri , based on material (holotype: UMMZ 131685; Fig. 108 View FIGURE 108 ) from “ 2.2 km (by road) W Patio de Aviación, Guerrero, Mexico, 2160 m.” Nieto Montes de Oca et al. (2013) placed A. adleri in the synonymy of A. liogaster as already suggested by Lieb (1981). Recently, Köhler et al. (2014) described a new species, Anolis peucephilus , from the pine forests of the Sierra Madre del Sur in Oaxaca; holotype SMF 96368 ( Fig. 109 View FIGURE 109 ) from “ca. 27 km on road N San Gabriel Mixtepec (16.19135°N, 97.09820°W, WGS84), 1325 m, Estado de Oaxaca, Mexico.”
The results of our molecular genetic analysis ( Fig. 110 View FIGURE 110 ) indicate several distinct genetic clusters in this complex, most of which can be assigned to species names with certainty (i.e., A. gadovii , A. liogaster , A. omiltemanus , and A. peucephilus ,). Specimens assigned to A. taylori form a cluster that is embedded within three clusters of specimens assigned to A. dunni . Relatively low genetic distances separate the four clusters in the dunni / taylori -clade. However, since A. taylori is readily diagnosable from A. dunni on the basis of male body coloration and male dewlap coloration, we recognize both nominal taxa as valid species. But we refrain from recognizing the three clades of dunni -like anoles as three species-level taxa because of the lack of morphological differentiation. Finally, we have examined the holotype of Anolis adleri Smith 1972 (UMMZ 131685; Fig. 108 View FIGURE 108 ) and agree with Nieto Montes de Oca et al. (2013) that it is conspecific with A. liogaster .










| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anolis nebulosus ( Wiegmann 1834 )
| Köhler, Gunther, Pérez, Raúl Gómez Trejo, Petersen, Claus Bo P., Méndez, Fausto R. & Cruz, De La 2014 |
Anolis schmidti
| Smith, H. M. 1939: 21 |
Dactyloa nebulosa
| Wiegmann, A. F. A. 1834: 47 |