
Archaebalaenoptera liesselensis, Bisconti & Munsterman & Fraaije, 2020
|
publication ID |
https://doi.org/ 10.7717/peerj.8315 |
|
publication LSID |
lsid:zoobank.org:pub:41E2B619-3E94-4299-956E-CAF7237A147F |
|
persistent identifier |
https://treatment.plazi.org/id/D6EF2A5E-B8C4-4D58-A141-633A72C177F2 |
|
taxon LSID |
lsid:zoobank.org:act:D6EF2A5E-B8C4-4D58-A141-633A72C177F2 |
|
treatment provided by |
Plazi |
|
scientific name |
Archaebalaenoptera liesselensis |
| status |
sp. nov. |
Archaebalaenoptera liesselensis sp. nov.
Diagnosis of species. Presently, autapomorphic characters are not observed in Archaebalaenoptera liesselensis ; this could be due to the fact that some of the anatomical districts preserved in the holotype cannot be investigated in the other Archaebalaenoptera species (e.g., periotic, endocranial cavity etc.). Archaebalaenoptera liesselensis differs from Archaebalaenoptera castriarquati in lacking the dorsally rounded dome at the anterior portion of the supraoccipital (Table S8 and Fig. S5 View Figure 5 ), it lacks the strong tubercles for the attachment of neck muscles on the lateral borders of the supraoccipital, and lacks narial processes of the interorbital region of the frontal. Archaebalaenoptera liesselensis differs from the undescribed new Archaebalaenoptera species (MHNL 1610; Bisconti et al., 2020, unpublished data; see Bisconti & Bosselaers, 2016 and Bisconti, Munsterman & Post, 2019 for an overview) in that it is larger (supraoccipital length of A. liesselensis is 310 mm and that of MHNL 1610 is 287 mm) and has a differently shaped interorbital region of the frontal (e.g., it does not have narial process and comparatively wider and shorter interorbital region of the frontal with respect to A. castriarquati ).
Holotype. Skull MAB002286 held by Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, The Netherlands .
Etymology. The species name, liesselensis , is from Liessel, the nearest village to the discovery site.
Horizon and locality. The skull MAB 002286 was dredged from a sand pit near the village of Liessel in Noord Brabant, The Netherlands (geographic co-ordinates of the site: 51 ◦ 25 44 N; 05 ◦ 49 47 E). The pit contains from 0 to 8 m Late Pleistocene sediments, from 8 to 13 m Early Pleistocene sediments, and from 13 to 45 m Late Miocene sediments (‘groenzanden - greensands’ with some wood and clay lenses) ( Peters, 2009). In The Netherlands, the latter sediments originate from the widespread marine Breda Formation, which, at this site, is deposited in a delta-front setting ( Peters, 2009). The holotype was (and to some extend still is) partly embedded in a dense glauconitic and sandy ‘greensand’ matrix from the Late Miocene (Tortonian; between 8.1 and 7.5 Ma; see below for full explanation) suggesting that it is from the Breda Formation.
RESULTS
Palynomorph analysis and age assessment
Palynofacies
The preservation of the palynomorph (dinoflagellate cyst and sporomorph) assemblage recovered from the greensands associated with the current balaenopterid fossil is moderate to good. The recovery of the palynomorphs however does not fully meet our standard level of counting at least 200 specimens (here 57%). The dinoflagellate cyst ratio is 42% (of the total sum of dinoflagellate cysts and sporomorphs; standard reference for percentages in this paper). Bisaccate pollen dominate the microflora (51%). Bisaccate pollen are formed by
gymnosperms (Gymnospermae). These are usually considered to flourish in a relatively dry environment as higher pine forests. However bisaccate pollen grains have a high buoyancy in both air and water, hence can easily be transported to distal marine settings, even open oceans, excluding any spores ( Abbink, 1998). The category spores are here present in relatively low percentages (7%). The highest numbers of marine dinocysts are reached by the taxa Spiniferites spp. and Operculodinium centrocarpum . The genus Spiniferites has a preferential orientation for open marine neritic conditions (e.g., Brinkhuis, 1994).
Operculodinium centrocarpum is a cosmopolitan, opportunistic taxon, hence influenced by multiple ecological factors Hennissen et al., 2017). The coastal marine taxon Lingulodinium machaerophorum is also well-represented. Remarkable are the relatively high values of heterotrophic genera like Barssidinium , Lejeunecysta and Selenopemphix . Heterotrophic dinoflagellate cysts are associated with nutrient-rich water. All indicators combined indicate nutrient-rich neritic conditions. No reworking is noted.
Age assessment
The chronostratigraphic range of the dinoflagellate cyst Impagidinium densiverrucosum indicates an age in the Late Miocene ( Von Daniels et al., 1990; Zevenboom, 1995).
The presence of Operculodinium piasecki fits with this dating. The most important chronostratigraphic marker in the assemblage is Hystrichosphaeropsis obscura . The Last Occurrence (LO) of this taxon defines the top of the Hystrichosphaeropsis obscura biozone of Denmark ( Dybkjaer & Piasecki, 2010) and the DN9 Zone of the Eastern USA and Germany ( De Verteuil & Norris, 1996;; Köthe, 2012). In the Netherlands the event is associated with the top of M14 Zone ( Munsterman & Brinkhuis, 2004), in the late Tortonian (excluding the latest part). Marker species for an early-mid Tortonian stage, or older, are all missing. Hence the most probable dating is in the Late Miocene, Zone M14, ca. 8.1–7.5 Ma Munsterman & Brinkhuis, 2004, recalibrated to ( Ogg, Ogg & Gradstein, 2016). This fits with earlier interpreted palynological results of the succession belonging to the Breda Formation, from which the specimen comes, in borehole B52C1978 (Hoogdonk).
Skull description
Overview and preservation
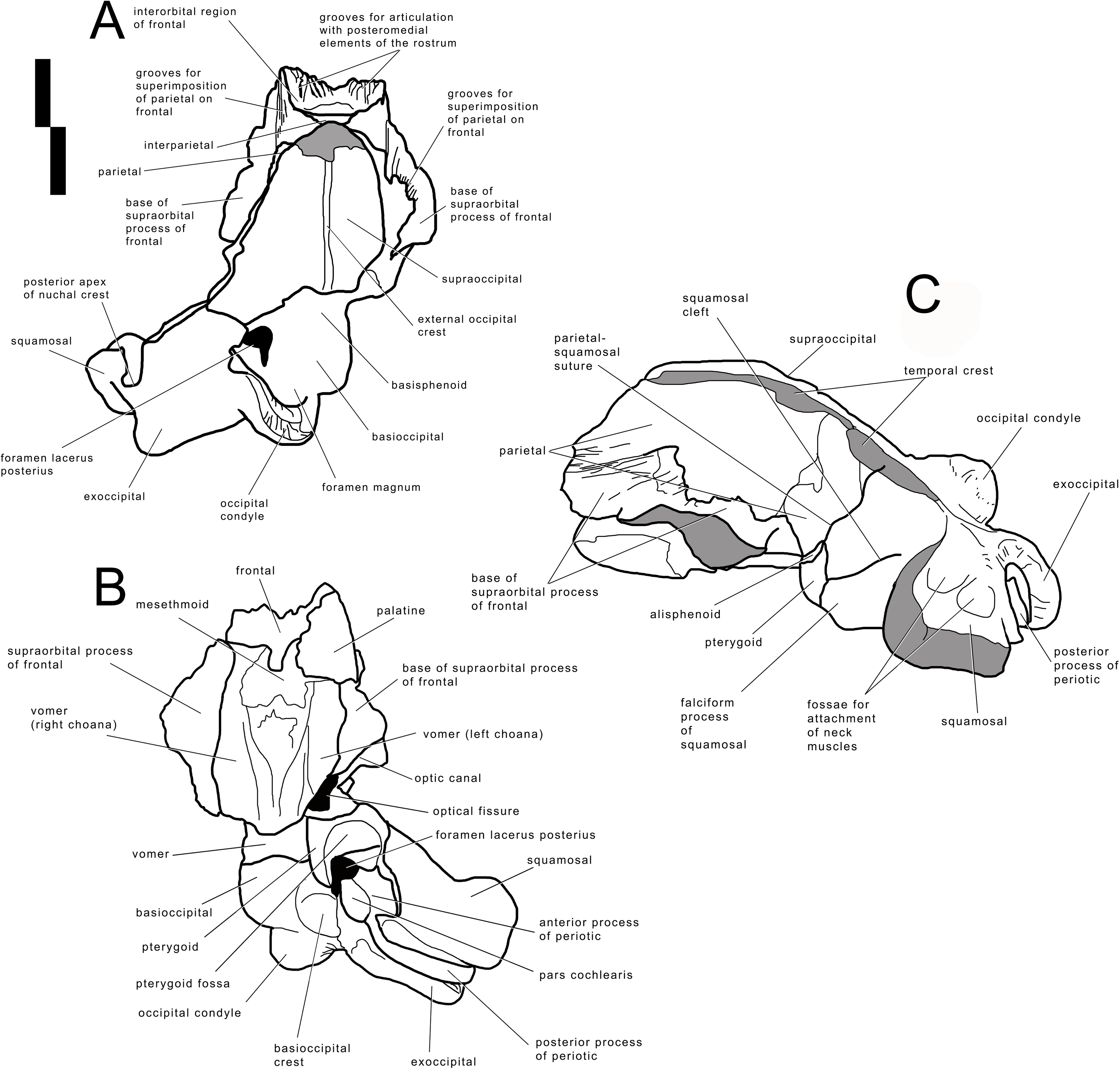
The skull of Archaebalaenoptera liesselensis lacks the rostrum, both supraorbital processes of the frontal, most of the squamosals and the tympanic bullae. One periotic is still in articulation. Most of the right side of the skull is broken and the endocranial cavity is exposed allowing the observation of the medial surface of the periotic. The remainder of the skull is well preserved. Standard views of the skull are presented in Figs. 2 View Figure 2 and 3 View Figure 3 . Measurements are provided in Tables S6 and S7 and discussed in the text below.
The lack of the lateral border of the supraoccipital is particularly problematic in the process of reconstructing the original skull shape. As shown in Fig. 2D View Figure 2 , in lateral view, the truncated border of the supraoccipital and of the parietal are evident. Observations of balaenopterid skulls in anterior view shows that the lateral borders of the supraoccipital do not reach the external rim of the temporal crest that is formed by the parietal ventrally and laterally (see, for instance, Marx & Kohno, 2016; Bisconti, Munsterman & Post, 2019). The lateral border of the supraoccipital is located slightly medially to the
external border of the skull at the level of the temporal crest. This means that a smaller portion of the supraoccipital was severed by post-mortem processes in the holotype skull of Archaebalaenoptera liesselensis . This consideration is further confirmed by the observation of the lateral view of the severed dorsolateral border of this skull where the larger severed component belongs to the parietal. Thus, we expect that the lateral extension of the supraoccipital was a little more developed than in the holotype skull but we do not think that such an extension was massively wider. We expect that only a few mm, maybe up to 10 mm, were lacking from the lateral border of the supraoccipital and this allows us to assess the general shape of the supraoccipital in dorsal view. Our description of the skull is also based upon this assumption.
Frontal
Interorbital region of the frontal and bases of supraorbital processes of the frontal are preserved ( Figs. 2 View Figure 2 and 3 View Figure 3 ). The interorbital region of the frontal shows articulation grooves for the attachment of the posterior end of the posteromedial elements of the rostrum. There are 6 grooves on the left side and 5 on the right side. It is difficult to determine which groove belongs to the articulation with the maxilla, which with the premaxilla
(if any) and which with the nasal. It is reasonable to suppose that most of the grooves belonged to the articulation with the maxilla suggesting that the posterior border of the ascending process of the maxilla was rounded and not squared as in the living species; this character is observed in all the Archaebalaenoptera species. Laterally to the grooves, the interorbital region of the frontal is a subtle stripe of bone that surrounds the grooves (and the posterior portion of the ascending process of the maxilla). The transverse diameter of the interorbital region of the frontal increases anteriorly. Along the longitudinal axis of the skull, the interorbital region of the frontal is located in between the grooves for the articulation with posteromedial elements of the rostrum but at a much lesser extension than in Archaebalaenoptera castriarquati . If the nasals would have been present between the ascending processes of maxillae and premaxillae, then, judging from the directions of the grooves for the articulation with the posteromedial elements of the rostrum, they should have had largely diverging lateral borders.
The supraorbital process of the frontal is abruptly depressed from the interorbital region of the frontal ( Fig. 4 View Figure 4 ). Judging from the bases of the supraorbital processes, these portions of the frontal were flat. The optic canal is located immediately anteriorly to the posterior border of the supraorbital process of the frontal ( Fig. S1 View Figure 1 ). There is no postorbital ridge. Grooves located on the dorsomedial surface of the supraorbital process of the frontal demonstrate that the parietal is spreading onto and covering the frontal along the depression lateral to the interorbital region of the frontal. The depressed portion of the interorbital region of the frontal has a round profile in cross section ( Fig. 4 View Figure 4 ). This character is typical and diagnostic of Archaebalaenoptera species. The supraorbital process of the frontal was dorsoventrally high (dorsoventral height along the posterior border
is 56 mm on the right side and 61 on the left side). A small foramen is observed on the right supraorbital process of the frontal; this foramen is prolonged into an anterolaterally directed groove.
Parietal
The anterior portion of the parietal projects anteriorly and interdigitates with the interorbital region of the frontal and the ascending process of the maxilla ( Figs. 2 View Figure 2 and 3 View Figure 3 ). In its anterior-most portion, the parietal is worn but the anterior point reached by the parietal is documented by the presence of grooves on the frontal. These grooves correspond to the surface of the overlapping of the parietal on the frontal. Anteriorly to the supraoccipital, the parietal is laterally convex along the dorsoventral axis of the skull forming an almost perfect half circle, as it is typical of the genus Archaebalaenoptera ( Fig. 4 View Figure 4 ). The parietal is externally concave at the base of the supraorbital process of the frontal. The anterior portion of the parietal projects medially and dorsally to surround the posterolateral corner of the interorbital region of the frontal; right and left parietals do not meet along the midline as they are separated by the interposition of an interparietal ( Fig. 3 View Figure 3 ). At the level of the maximum projection of the temporal crest, the parietal is strongly concave laterally along the dorsoventral axis of the skull ( Fig. 3 View Figure 3 ). The dorsal edge of the parietal is not preserved but appears to have protruded laterally following the lateral borders of the supraoccipital forming the temporal crest. Judging from the current state of preservation, the temporal crest was sufficiently developed to prevent the observation of the medial wall of the temporal fossa in dorsal view. The parietal-squamosal suture starts from the contact point of parietal and pterygoid immediately posterior to the alisphenoid exposure and proceeds dorsally and posteriorly along a sinuous path. A wide and triangular window is open on the medial wall of the temporal fossa 35 mm posteriorly to the supraorbital process of the frontal; this window is caused by damaging post-mortem processes and measures 35 mm in maximum diameter and 34 mm in minimum diameter.
Alisphenoid and temporal fossa
The alisphenoid exposure in the temporal fossa is limited to a small surface included between parietal, pterygoid and squamosal ( Fig. 3 View Figure 3 ). The dorsoventral diameter of the alisphenoid exposure is 20 mm and the anteroposterior diameter is 34.8 mm so that the bone is rectangular in shape. In the current preservation state, the superior portion of the falciform process of the squamosal terminates more posteriorly than the inferior portion resulting in a wider exposure of the dorsal portion of the pterygoid in the temporal fossa (the dorsoventral diameter of the pterygoid exposure is 62 mm and the anteroposterior diameter is 33 mm). Moreover, the ventral border of the parietal is partially eroded
so that a wider portion of the alisphenoid is exposed. The correct reconstruction of this part of the temporal fossa is provided in Fig. 3 View Figure 3 where it is possible to observe that the alisphenoid-parietal suture is dorsally concave; the alisphenoid-squamosal suture is limited to a point and the alisphenoid-pterygoid suture is convex. Based on this reconstruction, the alisphenoid exposure in the temporal fossa is limited to a short stripe of bone.
Squamosal
The parietal-squamosal suture begins from the posterodorsal corner of the alisphenoid and sinuously projects posterodorsally. Approaching the supraoccipital, the area including the parietal-squamosal suture becomes laterally convex along the anteroposterior axis. This area turns to be concave in close proximity to the posterior apex of the nuchal crest (sensu Mead & Fordyce, 2010; this portion is also known as lambdoid crest, see Mead & Fordyce, 2010, p. 34). The posterior development of the nuchal crest is rounded in cross-section and its posterior apex reaches a point located more anteriorly than the occipital condyles. The area included between the parietal-squamosal suture and the squamosal cleft is mostly flat along the dorsoventral and the anteroposterior axes; this area becomes convex in its more posterior portion. The squamosal cleft is straight and is 127 mm in length. It ends 62 mm anteroventrally from the posterior apex of the nuchal crest. The lateral surface of the squamosal shows two fossae for insertion of the neck muscles (very probably, the sternocephalicus muscle). These fossae are clearly separated by an anteroposterior crest.
In ventral view, the squamosal is a robust block characterized by a main axis forming nearly a right angle with the anteroposterior axis of the skull. External acoustic meatus and zygomatic process of the squamosal are not preserved entirely. Only the medial portion of the external acoustic meatus is present, it is 31 mm in anteroposterior diameter and shows a strong ventral concavity along its transverse development. Based on this observation, it is expected that the external acoustic meatus was long (probably 120+ mm) and anteroposteriorly narrow.
Pterygoid
In lateral view, only a small stripe of pterygoid is visible that corresponds to the dorsal lamina ( Fig. 3 View Figure 3 ). This portion is located anteriorly to the squamosal and ventrally to the alisphenoid. The pterygoid fossa is well preserved on the left side of the skull ( Fig. 2 View Figure 2 ). The palatal surface of the pterygoid fossa is anteriorly wide and its anterior and posterior borders are posteriorly concave. The posterior border of the palatal surface is obliquely oriented in the sense that it proceeds from an anterolateral point to a posteromedial point. The posterior border of the palatal surface forms the anterior border of the foramen lacerus posterius.
Supraoccipital
In dorsal view, the supraoccipital is roughly triangular with a remarkable transverse constriction located approximately at the middle of its anteroposterior length ( Figs. 2 View Figure 2 and 3 View Figure 3 ). The anterior portion is partially broken but the correct outline may be easily reconstructed by mirroring the contralateral part. As the lateral edges of the supraoccipital are broken, based on the thickness of the preserved borders, we estimate that the supraoccipital border protruded up to 10 mm more laterally in the anterior-most quarter of the supraoccipital, and up to 50 mm in the central portion. Based on these estimates, we reconstructed the extent of the transverse constriction of the supraoccipital and the shape of the temporal crest as shown in Fig. S5 View Figure 5 . The anterior border is rounded and transversely narrow. The lateral borders project laterally and posteriorly forming protruding temporal crests that prevent the observation of the medial wall of the temporal fossa in dorsal view. The maximum transverse constriction of the supraoccipital is located c. 200 mm from the anterior end and c. 220 mm from the posterior apex of the nuchal crest being approximately located at the middle of the anteroposterior length of the supraoccipital.
The dorsal surface of the supraoccipital is anteriorly worn for about 58 mm; more posteriorly, it shows the presence of an external occipital crest laterally bordered by a bilateral fossa that is anteroposteriorly and transversely concave. About 190 mm from the posterior apex of the nuchal crest, the supraoccipital inclination changes: in fact, the anterior portion of the supraoccipital is almost horizontal while the posterior portion is posteroventrally bent.
No tubercles for the attachment of neck muscles are observed.
Exoccipital
The wide exoccipital projects posteriorly and laterally ( Figs. 2 View Figure 2 and 3 View Figure 3 ). The posterolateral corner of the exoccipital reaches a point located slightly posteriorly to the posterior articular surface of the occipital condyle. The surface of the exoccipital is convex; the ventral border is straight in posterior view. The occipital condyle is wide and robust; its articular surface is dorsoventrally convex and transversely flat. The articular surface is mainly developed ventrally to the ventral border of the foramen magnum. Only a minimal portion of the foramen magnum is still preserved being bordered by the preserved part of the left occipital condyle, and this prevents us to describe this structure in detail.
Basioccipital
The ventral surface of the basioccipital is flat along the longitudinal axis of the skull ( Figs. 2 View Figure 2 and 3 View Figure 3 ); the surface becomes concave where the basioccipital crest are protruded. The basioccipital crest is tubercle-like and shows a ventrally-faced articular surface for the hyoid. The basioccipital crest forms the medial border of the foramen lacerus posterius; this border has a sinuous outline in ventral view. The occipital condyles and the basioccipital crests are separated by the interposition of a wide transverse groove (anteroposterior diameter of 40 mm). The jugular notch is wide, very concave and lacks any foramen. There is no condyloid foramen.
The endocranial cavity is exposed due to a large breakage of the right posterolateral side of the skull ( Fig. S2 View Figure 2 ). The endocranial face of the basioccipital and of the basisphenoid is mostly flat; the sella turcica is a slight and elliptical depression located slightly anteriorly to the anterior edge of the foramen lacerus posterius. The lateral wall of the endocranial cavity is almost uniformly concave with a notably transverse constriction at the level of the foramen magnum.
Palatine
A fragment of the left palatine is preserved in the anterior portion of the skull, ventrally ( Fig. 2 View Figure 2 ). It shows a convex external surface and is 16 mm in medial thickness. Its thickness decreases at mid-length where it is only eight mm. More posteriorly, the thickness increases again as it becomes 13 mm. The internal surface is highly concave and forms the internal border of the left choana. The choana is relatively wide with its maximum dorsoventral diameter of 62 mm, maximum transverse diameter of 44 mm and maximum length of 170 mm. Medially, the palatine stays on the vomer. Only the dorsal portion of the right choana is preserved. The choana has a dorsoventral diameter of 58 mm and a length of 200 mm.
Vomer
In anterior view, the vomer shows a rounded ventral border that corresponds to the posterior-most portion of the bone. The posterior keel is largely worn but is evident
along the more posterior portion (around 80 mm in length). Laterally to the keel, the lateral surface of the vomer is slightly concave along the dorsoventral axis. The posterior portion of the vomer is reduced to a very subtle bony lamina that is superimposed to the basioccipital-basisphenoid suture. The posterior border of the vomer is posteriorly concave.
Periotic
The left periotic is still firmly articulated with the skull and parts of its borders are still covered by hard matrix ( Fig. 5 View Figure 5 ). Measurements of the periotic are provided in Table S7.
Preparation was complete in the ventral surface but it is hard to get access to the medial surface where preparation is complicated and not complete. The posterior process is long and forms a right angle with the anterior process. It is anteroposteriorly compressed but robust. The distal portion is exposed laterally posterior to the postglenoid process of the squamosal and can be observed in the skull in lateral view ( Fig. 5 View Figure 5 ). The anterior border is in contact with the external acoustic meatus whose posterior border is formed by the posterior meatal ridge formed by the squamosal. This lamina is very subtle (anteroposterior thickness about 1.2 mm), ventrally concave and transversely elongated. The anteroposterior diameter of the external acoustic meatus decreases from the medial to the lateral portions (medial length, 24 mm; length at mid-length, 11 mm; lateral length, 11 mm).
The (broken) posterior pedicle for the tympanic bulla is located along the anteroventral border of the posterior process at about 122 mm from the posterolateral border and is anteroposteriorly compressed ( Fig. 5 View Figure 5 ). Its length is 16 mm. The same border forms the anterior edge of the facial canal that runs along the medial portion of the posterior process; the edge is not visible anymore starting from 81 mm from the lateral border of the posterior process.
Medially, the posterior process forms a curve with anterior concavity and is connected to the pars cochlearis. The length of the posterior process as a whole is c. 41 mm and the connecting neck to the pars cochlaris is particularly long. The groove for the facial nerve running along the posterior process is anteroposteriorly concave; it is 62 mm in length and 20 mm in width.
The anterior process is triangular in ventral view and its anterior end is located under the squamosal, just posteriorly to the posterior border of the pterygoid fossa. The posteromedial border of the anterior process shows a medial eminence with a convex medial border ( Fig. 5 View Figure 5 ). This eminence is called here Medial Eminence of the Anterior process (hereinafter: MEA). The peculiar shape of the MEA border gives the medial border of the anterior process a sinuous shape. A triangular and protruding lateral tuberosity is evident along the lateral border of the anterior process just laterally to the pars cochlearis. The (broken) anterior pedicle for the tympanic bulla is straight and narrow, and is located along the sharply-edged lateral border of the periotic. The anterior pedicle for the tympanic bulla is 15 mm in length and three mm in width. The ventral surface of the anterior process is flat-to-slightly concave near the lateral tuberosity; this surface is then anteroposteriorly convex approaching the medial border of the anterior process. A medial groove is interposed between the MEA and the anterior-most portion of the anterior process. This groove runs from a posteriormedial point to an anterolateral point and merges in the medial border of the anterior process.
The pars cochlearis is transversely protruded and is separated from the anterior process by an evident groove for the tensor tympani. The pars cochlearis is also ventrally protruding ( Fig. S3 View Figure 3 ). The distance between the superior border of the pars cochlearis and the ventral surface of the anterior process is reduced and, therefore, structures such as the distal opening of the facial canal and the oval window are dorsoventrally much smaller than expected based on comparative analysis of other living and fossil balaenopterid whales.
The caudal tympanic process is not preserved; however, a foramen is present to allow the passage of the facial nerve that merges into the groove for the facial nerve that is present along the posterior process of the periotic. The oval window is elliptical in shape (Table S7). The fossa for the stapedial muscle is highly compressed dorsoventrally and is not separated from the oval window by a crest. Only the posterior border of the round window is evident, the remainder and the entire posterior face of the pars cochlearis being invaded and covered by matrix.
The endocranial surface of the pars cochlearis cannot be observed in detail. In Fig. S4 View Figure 4 , it is possible to observe the endocranial opening of the facial canal located in the inferior-most part of the pars cochlearis itself. The suprameatal surface is highly concave but its dorsal border is still obliterated by matrix and cannot be distinguished from the endocranial face of the skull.
Endocranial cavity. The endocranial cavity is laterally concave and ventrally flat ( Fig. S2 View Figure 2 ). The lateral walls are formed by parietal, squamosal and alisphenoid. The alisphenoidparietal suture is highly interdigitated from the inside of the skull and terminates dorsally in a wide, triangular hole, and ventrally in the foramen lacerus posterius. The internal side of the alisphenoid is elongated and projects dorsally and anteriorly. It is not clear
if it contributes to the posterodorsal portion of the optic channel. The basioccipitalbasisphenoid is fused and its original position cannot be determined. The basisphenoidpresphenoid suture is still unfused. The internal surface of the basioccipital is transversely convex and anteroposteriorly flat. The sella turcica is a shallow and elliptical concavity in the endocranial surface of the basisphenoid. Posteriorly and laterally, the exoccipital is highly concave laterally to the foramen magnum.
COMPARISONS
Comparisons to other Archaebalaenoptera species
The new Archaebalaenoptera liesselensis differs from Archaebalaenoptera castriarquati and the MHNL1610 in a number of characters (see Table S8 and Fig. S5 View Figure 5 ). The supraoccipital of A. castriarquati shows a dorsal convexity at its anterior end ( Fig. S5 View Figure 5 ); we use the term ‘dome’ for this convexity in the following text. Instead of the dome, A. liesselensis shows a dorsally concave dorsal surface with an elongated and transversely narrow external occipital crest. The infraorbital region of the frontal of A. liesselensis lacks the elongated and triangular narial process observed in A. castriarquati as the frontonasal suture of the former is approximately straight (along the transverse axis) and that of the latter is obliquely oriented (from a posterolateral point to an anteromedial point being the anteromedial point located much more anteriorly than the posterolateral point). In A. castriarquati the anterior end of the parietal terminates at a protrusion located on the descending portion of the frontal just medial to the emergence of the supraorbital process of the frontal. Bisconti (2007) called this protrusion a ‘boss’ and noted that such a boss is also present in other balaenopterid species (e.g., Balaenoptera acutorostrata ). The boss is absent in A. liesselensis and the anterior end of the parietal is not anteriorly bounded by any detectable structure
located in the frontal bone. The posterolateral protrusion of the exoccipital of A. liesselensis is more marked than that observed in A. castriarquati as a distinctive concavity is present between the anterolateral corner of the exoccipital and the squamosal ( Fig. S5 View Figure 5 ) in A.
liesselensis but in A. castriarquati the continuity between the exoccipital and the squamosal is more evident in dorsal view. In A. castriarquati the nuchal crest does not overwhelm the medial wall of the temporal fossa but in A. liesselensis the temporal crest protrudes more laterally than in A. castriarquati and overwhelms the medial wall of the temporal fossa.
With respect to the other putative Archaebalaenoptera from Peru (MHNL1610), we can observe that the anterior end of the supraocciptal is almost squared in MHNL1610 and round in A. liesselensis , the development of the temporal crest is similar to that observed in A. castriarquati as the temporal fossa is completely exposed in dorsal view, and that the frontonasal suture is more similar to that observed in A. liesselensis . More details will be described and discussed elsewhere.
Comparisons with other balaenopterid taxa
The supraoccipital of Archaebalaenoptera is typical of this genus as, in dorsal view, it exhibits a marked transverse constriction, an elongated anterior portion and a short posterior portion. A transverse constriction is observed also in other balaenopterid taxa and in other mysticetes but it is characterized by different aspects in different taxa. In Protoroqualus, for instance, the transverse constriction is located anteriorly, very close
to the anterior end of the supraoccipital and the anterior portion of the supraoccipital converges toward the longitudinal axis of the skull forming an anteriorly pointed anterior border. In ‘Balaenoptera’ cortesii var. portisi the transverse constriction is much more marked and the anterior portion of the supraoccipital is transversely narrow and its anterior end is triangular; moreover, in this taxon, anteriorly to the transverse constriction, the lateral edges of the supraoccipital are more straight in dorsal view. In Nehalaennia devossi the supraoccipital shows a slight transverse constriction and the anterior end of the supraoccipital is wider and round. Interestingly, in ‘Balaenoptera’ bertae the transverse constriction is located in the anterior half of the supraoccipital (resembling Protororqualus and the portion anterior to the constriction is short and shows an approximately squared anterior end). In modern balaenopterids the transverse constriction is usually slight (see images in Bisconti, 2007 and True, 1904) and the anterior end is often squared ( Megaptera novaeangliae and B. acutorostrata are noteworthy examples in which the anterior end
of the supraoccipital may be also rounded; see True, 1904). Incakujira , Diunatans and ‘Balaenoptera’ siberi show no constriction at all.
The periotic of A. liesselensis shows a high suprameatal area with a strongly protruding dorsal surface. A strong dorsal protrusion is observed in Plesiobalaenoptera quarantellii ( Bisconti, 2010) . In A. liesselensis , the endocranial foramina are distributed in a subtle area located in the ventral-most portion of the pars cochlearis; this character is not observed in other balaenopterid taxa. The posterior process is elongated and narrow and shows a wide canal for the route of the facial nerve. In modern balaenopterid species the posterior process is flattened (Bisconti, 2001; ( Ekdale, Berta & Deméré, 2011) and the route of the facial nerve is not marked as in A. liesselensis . In Incakujira anillodefuego the pars cochlearis
is much more elongated with respect to the anteroposterior length of the anterior process Marx & Kohno, 2016). In Fragilicetus velponi and Diunatans luctoretemergo the periotic does not show the massive bulging of the dorsal surface and the anterior process is longer ( Bisconti & Bosselaers, 2016; Bisconti, Munsterman & Post, 2019).
A posterior protrusion of the exoccipital is observed in a number of fossil balaenopterid taxa, in eschrichtiids and cetotheriids. Bisconti & Bosselaers (2016) considered this character as a primitive state in the reconstruction of the morphological transformations of the balaenopterid skull. In particular, a protruded exoccipital is observed in Fragilicetus velponi , Archaebalaenoptera castriarquati and Eschrichtius robustus . Archaebalaenoptera liesselensis and A. castriarquati share this character with those taxa.
In A. liesselensis the alisphenoid is exposed in the medial wall of the temporal fossa for a small and rectangular spot between the parietal, squamosal and pterygoid. In the living balaenopterid species, Incakujira anillodefuego , Diunatans luctoretemergo, Norrisanima miocaena ( Leslie, Peredo & Pyenson, 2019) and Fragilicetus velponi , the alisphenoid is exposed as a very small area included between the parietal, squamosal and pterygoid. Exceptions to this pattern are the humpback whale, Megaptera novaeangliae , and Nehalaennia devossi in which the alisphenoid is not exposed at all (True, 1904; Bisconti, Munsterman & Post, 2019).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |