
Boninpagurus pilosipes (Stimpson, 1858) Stimpson, 1858
|
publication ID |
https://doi.org/ 10.5281/zenodo.202569 |
|
DOI |
https://doi.org/10.5281/zenodo.5667731 |
|
persistent identifier |
https://treatment.plazi.org/id/7E718793-8579-FFCD-FF56-FA93FB6FF99A |
|
treatment provided by |
Plazi |
|
scientific name |
Boninpagurus pilosipes (Stimpson, 1858) |
| status |
comb. nov. |
Boninpagurus pilosipes (Stimpson, 1858) View in CoL , n. comb.
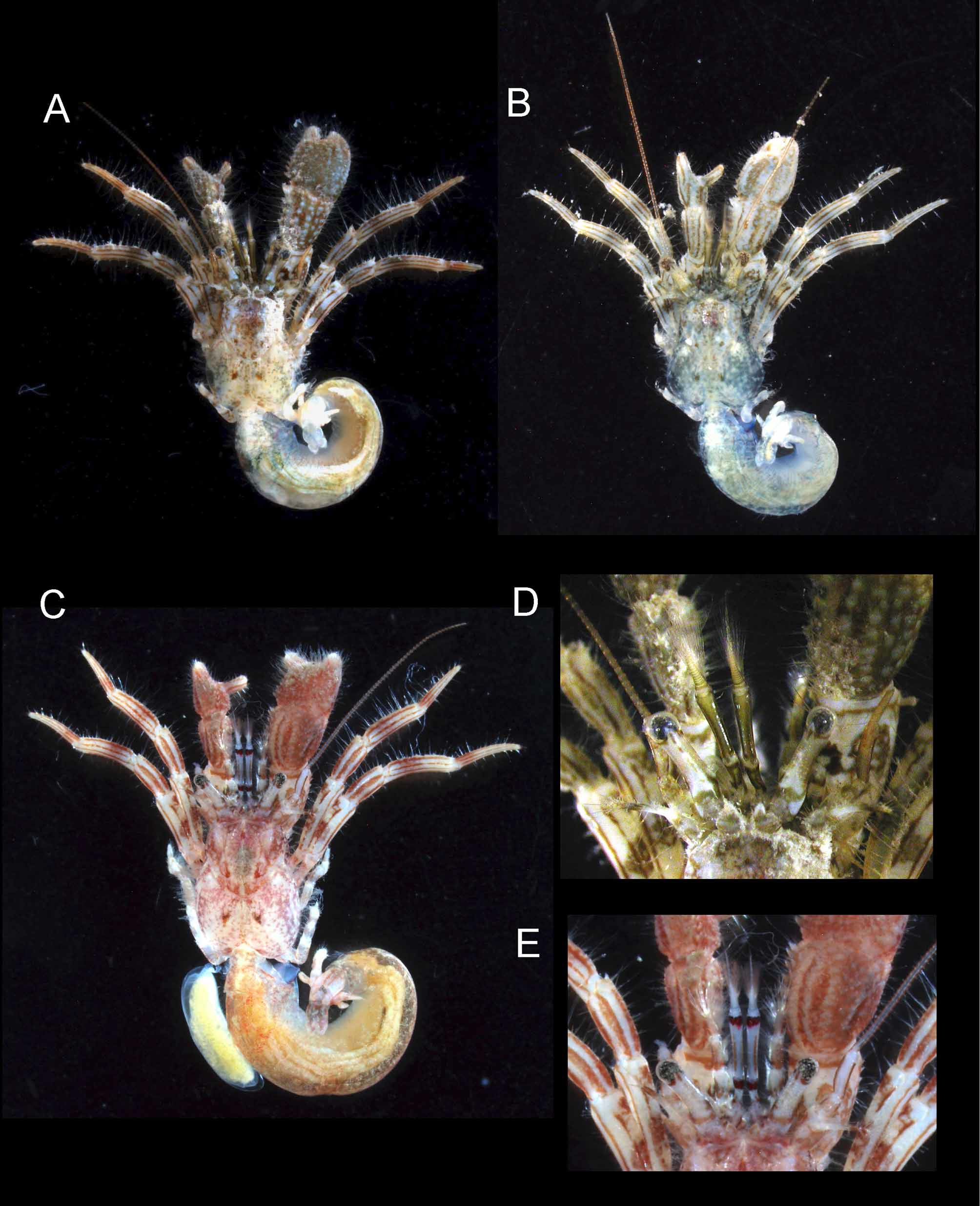
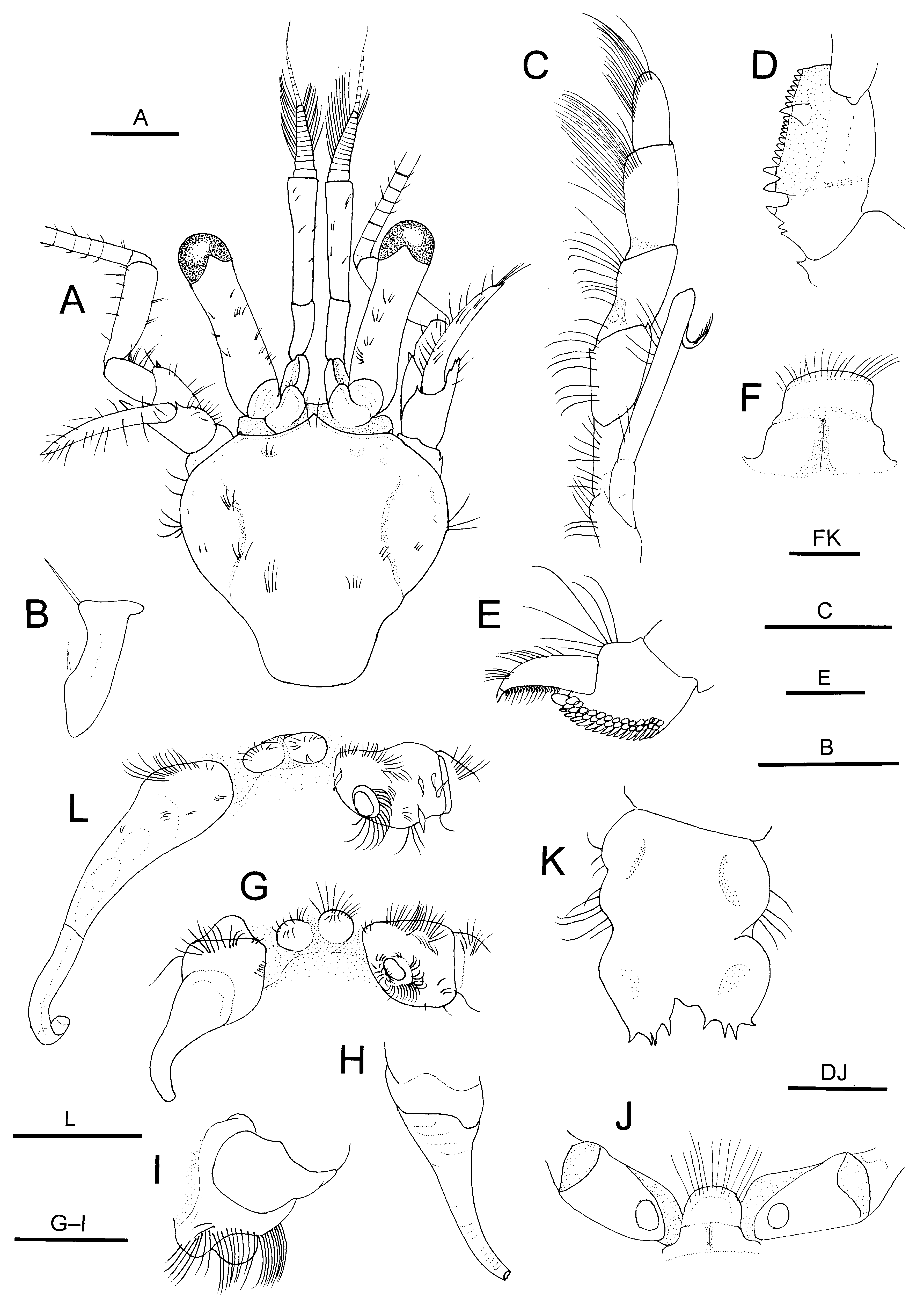
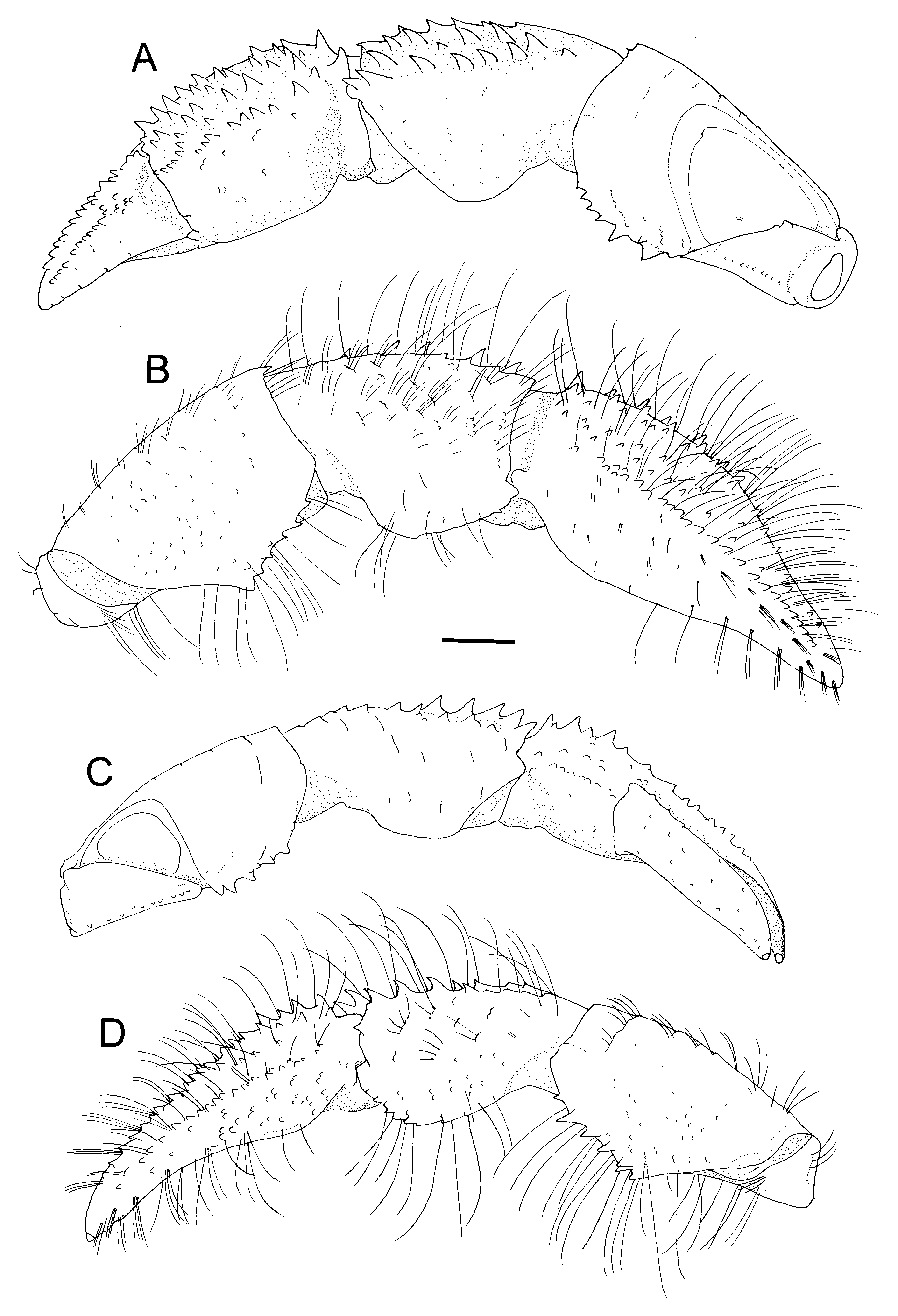
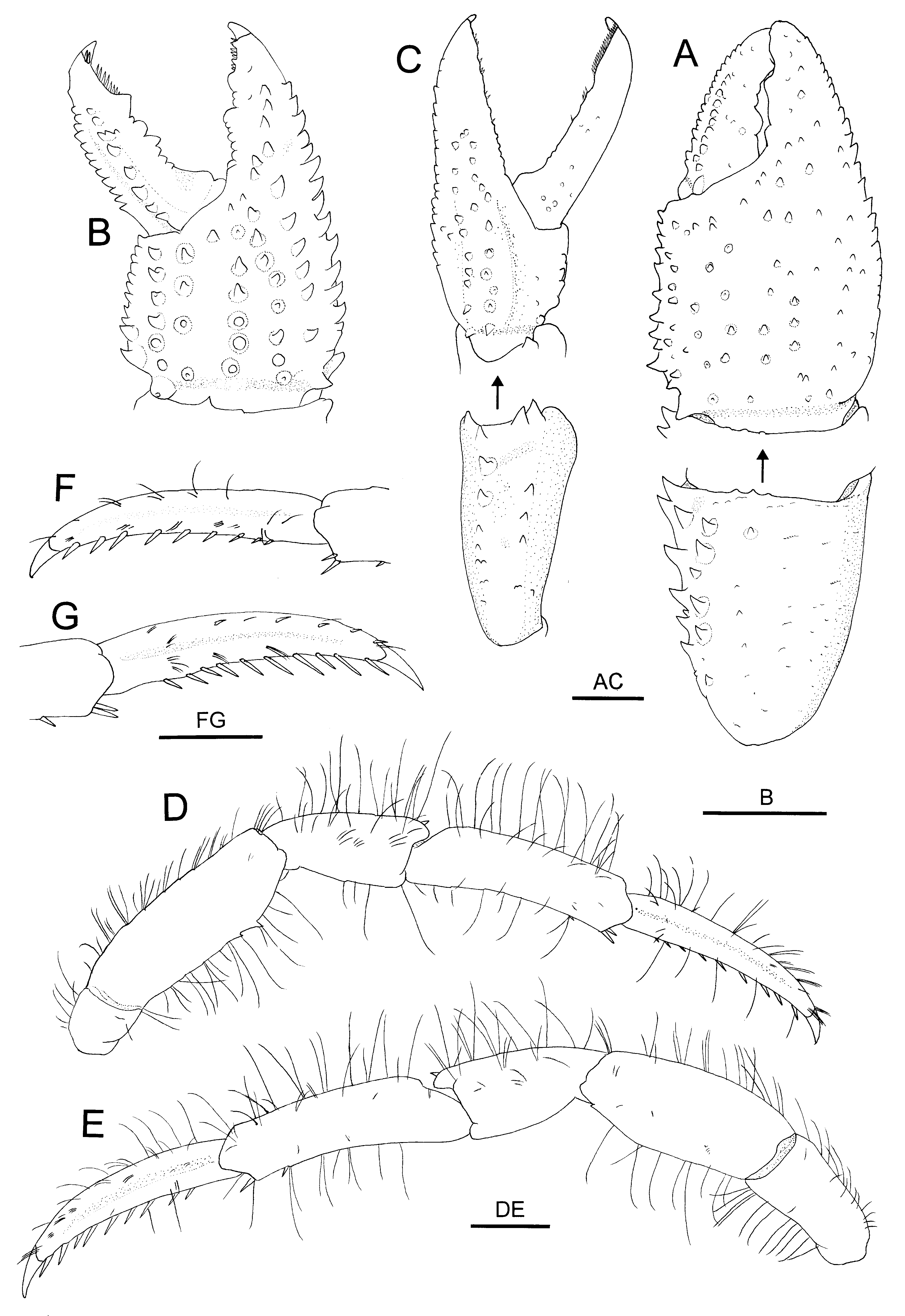
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Eupagurus pilosipes Stimpson, 1858: 249 View in CoL (87) [type locality: Loo Choo (= Okinawa Island)]; Alcock 1905: 177; Stimpson 1907: 133; Manning & Reed 2006: 285 (list).
Pagurus pilosipes View in CoL . — Oh 1983: 106 (in part), pl. 1, figs 3, 4, pl. 2, figs 1–5; Komai 2003: 117, figs 1–5, 24A; Hong et al. 2006: 254; McLaughlin et al. 2010: 34. [See Remarks]
Boninpagurus acanthocheles Asakura & Tachikawa, 2004: 158 View in CoL , figs 1–7 [type locality: Ogasawara Islands]; Asakura 2006: 145, figs 8, 9. McLaughlin et al. 2010: 27.
Boninpagurus View in CoL sp. — Okuno et al. 2006: 153; McLaughlin et al. 2010: 88, 89, fig. 12C.
Not Pagurus pilosipes View in CoL . — Miyake 1978: 91, fig. 34 (= mixture of seven species; see Komai, 2003); Miyake & Imafuku 1980: 60, pl. 2, fig. 5 (= Pagurus nigrivittatus Komai, 2003 View in CoL ); Miyake 1982: 132, pl. 44, fig. 5 (= P. nigrivittatus View in CoL ); Oh 1983: pl. 1, figs. 3, 4, pl. 3, figs. 1–5 (= P. nigrivittatus View in CoL ); Takeda 1986: 124, unnumbered fig. (= P. nigrivittatus View in CoL ); Asakura 1991: 798 (= P. nigrivittatus View in CoL ); Wang 1992: 61 (list); 1994: 570 (list); Asakura 1995: 363, pl. 97, fig. 9 (= P. nigrivittatus View in CoL ); Minemizu 2000: 148, unnumbered fig. (= P. nigrivittatus View in CoL ).
Material examined. Neotype: female (SL 2.5 mm), CBM-ZC 6383, Heshikiya, Katsuren Peninsula, Okinawa Island, intertidal, on small colony of Pocilopora sp., 23 June 1994, coll. T. Komai.
Japanese mainland: 1 female (SL 2.7 mm), CMNH-ZC 629, Igai-jima Islet, Kamogawa, 16 m, 29 June 2001, SCUBA diving, coll. J. Okuno; 1 male (SL 3.7 mm), 1 ovigerous female (SL 3.2 mm), CMNH-ZC 630, similar locality, 20 m, 16 November 2001, SCUBA diving, coll. J. Okuno; 1 ovigerous female (SL 2.7 mm), CMNH-ZC 631, same data; 1 male (SL 2.3 mm), CMNH-ZC, Ubara Islet, Katsuura, Boso Peninsula, 24 October 2001, SCUBA diving, coll. J. Okuno; 1 male (SL 2.7 mm), CMNH-ZC 1330, Awatone, off Kamogawa, 15–20 m, 18 December 2002, SCUBA diving, coll. H. Tachikawa; 1 male, CMNH-ZC 1331, same data as CMNH-ZC 1330; 1 male (SL 3.1 mm), CMNH-ZC 1346, Ashika-ne, off Kamogawa, Boso Peninsula, 26 m, 26 November 2000, SCUBA diving, coll. J. Okuno; 1 female (SL 2.5 mm), CMNH-ZC 1347, same data as CMNH-ZC 1346; 1 male (SL 3.4 mm), CMNH-ZC 1378, Hatto-ne, off Kamogawa, 15 m, 26 November 2000, SCUBA diving, coll. J. Okuno; 1 ovigerous female (SL 2.1 mm), CMNH-ZC 1615, Mankuro-ne, off Kamogawa, 20 m, 1 October 2003, SCUBA diving, coll. H. Tachikawa et al.; 1 female (SL 2.9 mm), CMNH-ZC 1729, Akino-hama, Izu-Oshima Island, Izu Islands, 15 m, 23 February 2004, SCUBA diving, coll. H. Arima; 1 ovigerous female (SL 1.8 mm), CMNH-ZC 1740, same locality, 15 m, 6 March 2004, SCUBA diving, coll. H. Arima; 1 male (SL 1.7 mm), CMNH-ZC 1743, same locality, 25 m, 13 March, 2004, SCUBA diving, coll. H. Arima; 1 male (SL 2.8 mm), CMNH-ZC 1837, Masaki, Shimizu, Suruga Bay, 8 m, 4 November 2005, SCUBA diving, coll. J. Okuno; 1 female (SL 2.0 mm), CMNH-ZC 1838, same data as CMNH-ZC 1837; 1 female (SL 2.0 mm), CMNH-ZC, Tatsunokuchi, Nagasaki, Kyushu, 2 m, 13 September 2003, SCUBA diving, coll. J. Okuno; 1 male (SL 2.0 mm), CMNH-ZC 1982, Mankuro-ne, off Kamogawa, Boso Peninsula, 18 m, 11 August 2005, SCUBA diving, coll. J. Okuno; 2 males (SL 2.8, 3.0 mm), CBM-ZC 10000, Sagami Bay, Hayama, Miura Peninsula, ca. 10 m, 23 September 2009, SCUBA diving, coll. I. Itou and S. Komai; 1 male (SL 2.2 mm), NTOU M01037 View Materials , 1 male (SL 2.1 mm), NTOU M01038 View Materials , same data as CBM-ZC 10000.
Okinawa Islands, Japan: 1 male (SL 1.9 mm), 2 females (SL 2.2, 2.4 mm), CBM-ZC 6384, same data as neotype; 2 males (SL 2.0, 2.0 mm), 1 female (SL 1.7 mm), CBM-ZC 10209, Heshikiya, Katsuren Peninsula, Okinawa Island, subtidal, dead coral, 17 July 2003, coll. M. Osawa; 1 male (SL 2.8 mm), CBM-ZC 10206, Kouri Fishing Port, Kouri Island, 2.0– 2.5 m, 25 February 2007, trap, coll. T. Komai; 1 ovigerous female (SL 3.3 mm), CBM-ZC 10275, Motobu Fishing Port, Motobu, Okinawa Island, 3–4 m, March 2010, trap, coll. T. Komai.
Ogasawara Islands, Japan: 2 males (SL 1.4, 1.7 mm), 2 females (SL 1.5, 1.5 mm), 2 juveniles (SL 1.2, 1.3 mm), CBM-ZC 10207, same locality, subtidal, coral rubble, 17 July 2009, coll. T. Komai; NTOU M 0 1039, 1 male (SL 1.4 mm), NTOU M01040 View Materials , 1 female (SL 1.4 mm), same data as CBM-ZC 10207.
Diagnosis. Shield ( Fig. 2 View FIGURE 2 A) as long as wide or slightly longer than wide; rostrum moderately broadly triangular, overreaching obtuse anterolateral projections; dorsal surface with distinct paragastric grooves. Ocular peduncles ( Fig. 2 View FIGURE 2 A) moderately slender, 0.7–0.8 times as long as shield; corneas not dilated; ocular acicles each with 1 submarginal terminal spine. Antennular peduncles ( Fig. 2 View FIGURE 2 A) overreaching distal corneal margins by 0.2–0.3 length of ultimate segment. Antennal peduncles ( Fig. 2 View FIGURE 2 A) reaching nearly to distal corneal margins; second segment with distolateral angle produced, reaching midlength of fourth segment, dorsomesial distal angle with small spine; antennal acicle slightly arcuate, reaching base of cornea. Third maxilliped ( Fig. 2 View FIGURE 2 C, D) with crista dentata consisting of closely spaced corneous teeth increasing in size proximally; carpus with small dorsodistal and ventrodistal spines. Chelipeds with scattered moderately long setae on dorsal surfaces of chela and carpus. Right cheliped ( Figs 3 View FIGURE 3 A, B, 4A, B) moderately stout; dactylus subequal in length to palm in males, distinctly longer than in females, dorsal surface with longitudinal row of small spines or spinulose tubercles adjacent to dorsomesial margin delimited by row of small spines; palm with 5–7 irregular rows of small spines or spiniform tubercles on dorsal surface, dorsolateral and dorsomesial margins each delimited by row of small spines; carpus with row of strong spines on dorsomesial margin and additional row of spines adjacent to dorsomesial margin, dorsolateral face rounded; merus with row of spines on ventrolateral and ventrodistal margins. Left cheliped ( Figs 3 View FIGURE 3 C, D, 4C) moderately slender; dactylus about twice as long as palm, without distinct spines on surfaces; palm with 2 or 3 rows of small to moderately small spines extending onto fixed finger on dorsal surface, dorsal midline weakly elevated, dorsolateral margin delimited by row of small spines, dorsomesial margin with tiny blunt tubercles; carpus weakly widened distally, with row of small spines on dorsolateral and dorsomesial margins; merus with row of small spines on ventrolateral and ventromesial margins. Ambulatory legs ( Fig. 4 View FIGURE 4 D, E) with tufts of stiff setae on dorsal and ventral margins; dactyli 1.0–1.1 times longer than propodi, each with 6–11 corneous spines on ventral margin, mesial face unarmed (second) ( Fig. 4 View FIGURE 4 F) or armed with row of spinules adjacent to dorsal margin (third) ( Fig. 4 View FIGURE 4 G); propodi each with 1 or 2 corneous spinules on ventrodistal margin and 1 or 2 spinules on ventral margin; carpi only with dorsodistal spine; meri each with few small spines on ventral margin (second) or unarmed (third). Coxa of right fifth pereopod of male with sexual tube of medium length (2–4 times coxal length), straight or slightly curved, directed laterally or posterolaterally, gradually tapering distally to truncate tip, proximal half opaque, and remaining distal portion transparent. Sixth thoracic sternite ( Fig. 2 View FIGURE 2 F, J) with subrectangular anterior lobe. Eighth thoracic sternite ( Fig. 2 View FIGURE 2 G, L) consisting of 2 rounded lobes. Telson ( Fig. 2 View FIGURE 2 K) with terminal margins slightly to somewhat oblique, each with 4 or 5 moderately large spines.
Coloration in life ( Fig. 1 View FIGURE 1 A, B, D). General background color gray- or green-brown. Shield mottled grayish or greenish brown, paler laterally, with 2 or 3 paired spots. Posterior carapace also mottled brown, with conspicuous pair of spots on either side of posteromedian plate. Ocular peduncles generally light brown, proximally with dark brown transverse band; proximal half of dorsal surface grayish. Antennular peduncles generally grayish or greenish brown; ultimate segment with 2 longitudinal dark brown stripes on dorsal surface; median part of dorsal surface of penultimate segment paler; articulation between segments whitish; dorsal flagellum pale brown. Antennal peduncles generally greenish or grayish brown; fifth segment with brown longitudinal stripes laterally and mesially; antennal flagellum entirely light brown, without particular pattern. Right cheliped with tips of fingers whitish; palm generally brown or grayish brown, with rather obscure longitudinal stripes laterally and mesially; carpus apparently with longitudinal stripes of brown and light gray on dorsal surface, latter consisting of longitudinal row of light gray spots; merus with subdistal, narrow, transverse brown band and irregular dark brown patches or blotches. Left cheliped with similar pattern to right cheliped, but longitudinal stripes on palm, carpus and merus more clearly delineated.
Distribution. Okinawa Islands (Katsuren and Motobu Peninsulas on Okinawa Island, and Kouri Port on Kouri Island), Chichi-jima Island of the Ogasawara Islands, Izu-oshima Island of the Izu Islands, mainland of Japan (Boso Peninsula on Honshu southward to Nagasaki on Kyushu), and southern part of Korea); intertidal to 25 m depth.
Remarks. Asakura & Tachikawa (2004) did not compare their new taxon B. acanthocheles with P. pi lo s ip e s in spite of close similarities, perhaps because the latter species was considered to lack sexual tube development at the time. The present newly collected male specimens from Okinawa Islands show sexual tube development, closely agreeing with that seen in B. acanthocheles . In this study, we examined specimens from various localities in Japan, including those from Okinawa and Ogasawara Islands, representing the type localities of the two taxa. As mentioned before, two morphs separated by the color of the antenular peduncle are recognized. Nevertheless, we could not find a single morphological character or a suite of characters that will separate one species from another. Comparison of partial sequence of the barcoding gene COI (657 bp) shows that there are only 0.6–1.5% genetic divergences amongst the specimens from the Japanese mainland, and Okinawa and Ogasawara Islands, while specimens from the same locality have genetic divergences of 0.5–1.4% ( Table 2 View TABLE 2 ). Such a low genetic divergence is generally considered as an intraspecific variation in decapod crustaceans (e.g. Khamnamtong et al. 2009; Rhyne et al. 2009; Silva et al. 2010; Yang et al. 2010). Thus, our initial suspicion that two species might be represented in the present collections was not supported by either morphological or molecular comparison. Consequently, we propose to synonymize B. acanthocheles under B. pilosipes , and refer all examined specimens to B. pilosipes .
Komai (2003) designated a neotype for Pagurus pilosipes in the interests of nomenclatural stability, because it was considered that the type was destroyed in the Great Chicago Fire of 1871 ( Evans 1967). Subsequently, Manning & Reed (2006) published a list of crustaceans that were shipped by W. Stimpson in 1859 to the Zoological Museum in Copenhagen (ZMC). In the list, Pagurus pilosipes is mentioned (as Eupagurus ), but no specimen was located in the collection of the ZMC. Therefore, the action by Komai (2003) is still warranted.
| NTOU |
Institute of Marine Biology, National Taiwan Ocean University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Boninpagurus pilosipes (Stimpson, 1858)
| Komai, Tomoyuki, Yang, Chien-Hui, Okuno, Junji & Chan, Tin-Yam 2011 |
Boninpagurus
| McLaughlin 2010: 88 |
| Okuno 2006: 153 |
Boninpagurus acanthocheles
| McLaughlin 2010: 27 |
| Asakura 2006: 145 |
| Asakura 2004: 158 |
Pagurus pilosipes
| McLaughlin 2010: 34 |
| Hong 2006: 254 |
| Komai 2003: 117 |
| Oh 1983: 106 |
Pagurus pilosipes
| Minemizu 2000: 148 |
| Asakura 1995: 363 |
| Wang 1992: 61 |
| Asakura 1991: 798 |
| Takeda 1986: 124 |
| Miyake 1982: 132 |
| Miyake 1980: 60 |
| Miyake 1978: 91 |
Eupagurus pilosipes
| Manning 2006: 285 |
| Stimpson 1907: 133 |
| Alcock 1905: 177 |