
Paranaitis
|
publication ID |
https://doi.org/10.1046/j.1096-3642.2003.00069.x |
|
persistent identifier |
https://treatment.plazi.org/id/793A8787-5C14-6038-FC02-FF1DFB5ACDD3 |
|
treatment provided by |
Carolina (2021-08-31 19:56:24, last updated by Plazi 2023-11-06 11:53:00) |
|
scientific name |
Paranaitis |
| status |
|
REVISION OF PARANAITIS View in CoL 407
Pygidial cirri cylindrical with rounded end, about 2.5 times as long as wide ( Fig. 23A View Figure 23 ). Pygidial papilla not observed.
Habitat
Shell sand, 55–59 m.
Distribution
Known only from east Japan.
Remarks
The present new species shares a deep ligula, presence of eyes, presence of chaetae on segment 2, chaetal morphology, and dorsally elongated dorsal cirrophores, with P. polynoides . It differs in having the proximal part of proboscis covered by large, as well as small, chitinous papillae (small only in P. polynoides ). The pigmentation on the dorsal cirri also differs from P. polynoides , having a few to several spots, which are frequently fused to each other and form irregular asterisk-shaped marks, as compared to single rounded ones in P. polynoides .
Etymology
This species is named for Mr Koichi Morita of Otsuchi Marine Research Center, in recognition of his vast knowledge of marine life.
PARANAITIS POLYNOIDES ( MOORE, 1909) View in CoL
( FIGS 24–26 View Figure 24 View Figure 25 View Figure 26 , 37 View Figure 37 )
Anaitis polynoides Moore, 1909: 339–342 View in CoL , pl. 16, figs 19–21; Berkeley, 1924: 287; Zachs, 1933: 127 (misspelled as Anaites polynoides ).
Paranaitis polynoides: Hartman, 1936: 117 View in CoL ; 1959: 158; 1968: 291, figs 1–3; Hartman & Reish, 1950; 12; Uschakov, 1972: 1141–142, pl. 7, figs 1–4; Blake, 1994: 164–165, fig. 4.22.
Phyllodoce (Anaitis) polynoides: Annenkova, 1937: 156 .
Not Paranaitis polynoides: Fauchald, 1972: 52 View in CoL (= Paranaitis sp. ); Gathof, 1984: 19-21 to 19-23, figs 19-17, 18a–e (= Paranaitis gardineri View in CoL ).
Material examined
Holotype (USNM-17267), Monterey Bay, California, 83 m; 1 specimen (USNM-123242), off California, 38∞16.14¢N, 123∞24.18¢W, 184 m; 1 specimen (USNM- 26839), off Apple Cove, Puget sound, Washington, shell, fine gravel and mud, 27 m; 1 specimen (USNM-32461), South Gedney Island , Puget Sound , Washington, 108 m; 1 specimen (ZIHU-2016), Palos Verdes shelf, Santa Monica Bay , California, 33∞43.14¢N, 118∞24.66¢W, olive green silt, 307 m, 15 January 1997, coll. F.P.
Description
Holotype complete specimen, 40 mm long, 3.4 mm wide, for 88 segments; see Figure 24 View Figure 24 for measurements of other specimens. Preserved specimen white to yellow, with reddish brown pigmentation dorsally from c. segment 7. Single large rounded reddish brown spot present on inside of dorsal cirri. Body stout, dorsoventrally flattened, anteriorly and posteriorly tapered. Prostomium rounded, wider than long, with distinct deep ligula ( Fig. 24A View Figure 24 ). Paired antennae and palps conical, directed laterally. Eyes large, rounded, with lenses, situated at posterior margin of prostomium, often partly covered by segment 1. Nuchal papilla small, rounded, situated posteriorly in ligula. Nuchal organs small, rounded, ventro-laterally situated. Proboscis with distinct division separating proximal and distal parts ( Fig. 24B View Figure 24 ). Proximal part entirely covered by diffusely distributed minute and conical chitinous papillae; size and shape of papillae similar on whole proximal part. Proximal-most part with 1–2 large conical papillae each side. Distal part with six indistinct longitudinal rows of tubercles, conical papillae absent. Terminal ring with c. 20 poorly delineated papillae. Paired large papillae present laterally inside ring.
Segments 1 and 2 fused. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered ( Fig. 25A–C View Figure 25 ). Cirri of segment 1 short, reaching about anterior margin of segment 4. Dorsal cirri of segment 2 reaching c. segment 7. Ventral cirri of segment 2 reaching c. segment 5. Dorsal cirri of segment 3 reaching c. segment 8. Segment 2 with small neuropodial lobes fusing to ventral cirrophores, with ventral aciculae and c. three chaetae. Segment 3 with neuropodia with ventral aciculae and c. eight chaetae, and ventral cirri of similar size and shape as following segments. Dorsal aciculae of segment 2 and 3 absent.
Dorsal cirri of median segments reniform, symmetrical, c. twice as wide as long. Dorsal cirrophores asymmetrical, with narrow extensions on dorsal side of cirri ( Fig. 25F View Figure 25 ). Neuropodium with supra-acicular lobes longer than subacicular lobes, with c. 20 chaetae. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, with single large main tooth on anterior side ( Fig. 37F View Figure 37 ). Ventral cirri oval with rounded ends ( Fig. 25G View Figure 25 ). Pygidial cirri cylindrical with rounded ends, 3–3.5 times as long as wide ( Fig. 25H View Figure 25 ). Pygidial papilla present.
Habitat
Sand, mud, and rocks, intertidally to 286 m.
Distribution
Pacific coast of Canada ( Berkeley, 1924) to southern California.
Remarks
Paranaitis polynoides differs from other Paranaitis in the character combination wide reniform dorsal cirri with dorsally elongated dorsal cirrophores, and proboscis with distinct division separating proximal and distal parts.
We consider Uschakov’s (1972) record from the north-west Pacific as doubtful, based on his description of lateral rows of papillae on the proximal part of proboscis.
Paranaitis polynoides has been reported both from the Atlantic and the Pacific coasts of USA and Canada. We examined all specimens (13 lots) identified as P. polynoides and deposited in USNM, and found that only the Pacific specimens actually conform to P. polynoides . The Atlantic specimens, including Gathof ’s material (USNM-56127), were re-identified as P. gardineri .
Fauchald (1972) reported this species from deep waters (2401–2480 m) off California. The specimen ( LACM Vel. 7231) has a retracted proboscis, but differs from P. polynoides in having long oval dorsal cirri and symmetrical dorsal cirrophore, and probably belongs to an undescribed species of Paranaitis . However, in the absence of information on a series of characters, we prefer to leave this single specimen undescribed.
PARANAITIS PUMILA SP. NOV.
( FIGS 27–29 View Figure 27 View Figure 28 View Figure 29 , 37 View Figure 37 )
Material examined
Holotype (ZIHU-1378), 21 paratypes (ZIHU-1379), 18 paratypes ( MNHN POLY TYPE 1379 ), Otsuchi Bay ,
Iwate, Honshu , Japan, 39∞23.1¢N, 141∞58.8¢E, shell sand, 67 m, 11 August 1998 , coll. T.K. 11 specimens (ZIHU-1968), Otsuchi Bay , 39∞21.0¢N 141∞58.5¢E, shell sand, 58.7 m, 7 May 1997 , coll. T.K.; 17 specimens (ZIHU-1969), Otsuchi Bay , 39∞20.5¢N, 141∞57.5¢E, shell sand, 45.3 m, 25 May 1998 , coll. T.K.; 60 specimens (ZIHU-1970), Otsuchi Bay , 39∞23.2¢N, 141∞58.8¢E, shell sand, 62.5 m, 26 May 1998 , coll. T.K.; 24 specimens (ZIHU-1971), Otsuchi Bay , 39∞20.8¢N, 141∞58.4¢E, shell sand, 53 m, 10 August 1998 , coll. T.K.; 10 specimens (ZIHU-1972), Otsuchi Bay , 39∞21.0¢N, 141∞58.4¢E, shell sand, 59 m, 10 August 1998 , coll. T.K.; 1 specimen (ZIHU-1973), Otsuchi Bay , 39∞22.0¢N, 142∞00.8¢E, sand, 98.9 m, 27 September 2000 , coll. T.K.; 12 specimens (ZIHU-1974), Otsuchi Bay , 39∞23.3¢N, 141∞59.7¢E, sand, 80.6 m, 27 September 2000 , coll. T.K.; 28 specimens (ZIHU-1975), Otsuchi Bay , 39∞23.4¢N, 141∞59.0¢E, shell sand, 64.0 m, 27 September 2000 , coll. T.K.; 1 specimen (ZIHU-1976), Otsuchi Bay , 39∞21.4¢N, 141∞59.1¢E, shell sand, 70.3 m, 27 September 2000 , coll. T.K.; 1 specimen (CMNH-ZW-832), Otsuchi Bay , 39∞ 22.16 N, 141∞ 59.97E, 87 m, 14 October 1996 GoogleMaps .
Median part of dorsum pale yellow. Yellow pigmentation lost in preserved specimens, but longitudinal lines retained. Eyes brown. Body of uniform width. Prostomium anteriorly rounded, wider than long, posteriorly with distinct deep ligula ( Fig. 27A View Figure 27 ). Paired antennae and palps narrow, anteriorly directed. Eyes large, rounded, with lenses, situated at posterior margin of prostomium, often partly covered by segment 1. Nuchal papilla rounded, situated posteriorly in ligula. Nuchal organs oval, ventro-laterally situated. Proboscis short, not divided into proximal and distal parts, lacking papillae or tubercles ( Fig. 27B, C View Figure 27 ). Terminal ring with c. 17 papillae. Paired large papillae present laterally inside ring.
Segments 1 and 2 fused. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered ( Fig. 28A–C View Figure 28 ). Cirri of segment 1 reaching c. segment 5. Dorsal cirri of segments 2 and 3 reaching c. segment 7. Ventral cirri of segment 2 reaching c. segment 5. Segment 2 with well-developed neuropodia near cirrophores of ventral cirri, with ventral aciculae and c. six chaetae. Segment 3 with neuropodial lobes with ventral aciculae and c. six chaetae, with ventral cirri of almost same shape and size as on following ones. Dorsal aciculae of segments 2 and 3 absent.
Dorsal cirri of median segments rounded cordate, symmetrical, about as long as wide ( Fig. 28D–H View Figure 28 ). Dorsal cirrophores small, dorsally indistinct. Neuropodium with supra-acicular lobes longer than subacicular lobes, with 12–15 chaetae. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, with single large main tooth on anterior side ( Fig. 37G View Figure 37 ). Ventral cirri oval, slightly shorter than neuropodia. Pygidial cirri oval 1.5–2 times as long as wide ( Fig. 27E View Figure 27 ). Pygidial papilla present.
Description
Holotype complete ovigerous female, 8.5 mm long, 0.8 mm wide, for 52 segments; see Figure 29 View Figure 29 for measurements of other specimens. Live animals white with reddish brown pigmentation on each side of dorsum, forming two longitudinal lines ( Fig. 27D View Figure 27 ). Habitat
Shell sand, 45–67 m.
Distribution
Known only from Otsuchi Bay, Japan.
Biology
Mature males and females collected in August at Otsuchi Bay.
Remarks
Paranaitis pumila sp. nov. differs from other Paranaitis in the character combination presence of eyes, anteriorly directed narrow paired antennae and palps, deep ligula, neuropodial lobes with chaetae on segment 2, and undivided proboscis lacking chitinous papillae.
Etymology
The new species is named for its small body size, ‘ pumila ’ being Latin for dwarf.
PARANAITIS SPECIOSA ( WEBSTER, 1879) View in CoL
( FIGS 30–33 View Figure 30 View Figure 31 View Figure 32 View Figure 33 , 37 View Figure 37 )
Anaitis speciosa Webster, 1879: 131–132 View in CoL , pl. 1, figs 8, 9; Webster & Benedict, 1884: 702, 1887: 710.
Paranaitis speciosa: Hartman, 1959: 158 View in CoL ; Pettibone, 1963: 75–77, fig. 17a; Day, 1973: 22; Gathof, 1984: 19-23 to 19-25, figs 19-19, 20a–e.
Material examined
USA: 4 syntypes (USNM-380), Great Egg Harbor, New Jersey ; 4 specimens (USNM-485), Provincetown, Massachusetts ; 3 specimens (USNM-26972), Great Egg Harbor, New Jersey ; 1 specimen (USNM-75588), Gulf of Mexico, Alabama, off Mobile Bay , 29∞43.29¢N, 87∞54.30¢W, 35 m ; 1 specimen (USNM-89977), Gulf of Mexico, Mississippi, Mississippi Sound , 30∞04.54¢N, 88∞49.54¢W, 12.2 m ; 1 specimen, Beaufort , North Carolina, shell sand, 12 m, 4 May 1972, coll. M.E. Petersen ; 7 specimens, Bogue Banks, Morehead , North Carolina, 28 February 1965, coll. M.E. Petersen. JAPAN: 1 specimen (ZIHU-1920), Otsuchi Bay, Iwate, Honshu , 39∞20.5¢N, 141∞58.4¢E, sandy mud, 41 m, 25 May 1998, coll. T.K .; 1 specimen (ZIHU-1921), 39∞20.8¢N, 141∞57.9¢E, Otsuchi Bay , sand, 48 m, 10 August 1998, coll. T.K .
Description
Largest syntype complete, whole mounted specimen, 4.5 mm long, 1.0 mm wide, for 35 segments larger specimen with regenerated caudal end 12.0 mm long, 1.6 mm wide, for 51 segments (ZIHU-1920). See Figure 33 View Figure 33 for measurements of other specimens. Live animals reddish yellow to very dark brown with darker transverse band on segments 8 and 9 ( Webster, 1879; new observations on Japanese specimens). Preserved specimens pale.
Body stout, dorso-ventrally flattened, anteriorly and posteriorly tapered ( Fig. 30A View Figure 30 ). Prostomium rounded, wider than long. Indistinct but deep ligula present in syntypes ( Fig. 30B View Figure 30 ); absent in larger specimens ( Figs 31A View Figure 31 , 32A View Figure 32 , ZIHU-1920, 1921). Paired antennae short and narrow, anteriorly to postero-laterally directed, c. one-third as long as wide of prostomium. Palps similar to paired antennae in length, slightly wider. Eyes medium sized, rounded, with lenses, situated at posterior margin of prostomium. Nuchal papilla indistinct, small, rounded, inserted posteriorly in ligula in syntypes; not detected in larger specimens (specimens from North Carolina, ZIHU-1920, 1921). Nuchal organs elongated oval, laterally situated
Segments 1 and 2 fused. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered. Cirri of segment 1 short, reaching about anterior margin of segment 4. Dorsal and ventral cirri of segment 2 reaching about middle of segment 5. Dorsal cirri of segment 3 reaching about middle of segment 6. Chaetae absent on segment 2. Distribution of aciculae in segments 2 and 3 unknown.
Dorsal cirri of median segments broad cordiform, with rounded ends ( Figs 30C,D View Figure 30 , 31C,D View Figure 31 , 32D,E View Figure 32 ). Dorsal cirrophores short and indistinct. Neuropodium with supra-acicular lobes longer than subacicular lobes, with c. 50 chaetae. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, with single large main tooth on anterior side ( Figs 37H,I View Figure 37 ). Ventral cirri reniform with rounded ends, shorter than neuropodial lobes. Pygidial cirri oval with rounded ends. Pygidial papilla present.
Habitat
Found in Mytilus beds, in tubes of Diopatra cuprea , in sand, mud, silt, clay and shell bottom, intertidally to 185 m.
Distribution
Maine to North Carolina; Gulf of Mexico ( Gathof, 1984).
( Fig. 31B View Figure 31 ). Proboscis dorsally covered with minute rounded papillae, with single lateral rows of large rounded papillae each side ( Figs 32B,C View Figure 32 ). Terminal ring with about 30 papillae. Paired large lateral papillae present inside ring.
Remarks
Paranaitis speciosa differs from other Paranaitis in the character combination indistinct nuchal papilla, proboscis with lateral row of large papillae on each side, and rostrum of chaetal shaft with single large main tooth. It is similar to P. wahlbergi in the indistinct nuchal papillae (at least in large specimens), and in the parapodia with small indistinct dorsal cirrophores, but differs in the morphology of the rostrum of the chaetal shaft. These two species can also be distinguished by pigmentation: live European P. wahlbergi have a clear red dorsum, whereas P. speciosa has a dark green, yellow, or yellow brown dorsum with a transverse dark dorsal band on segments 8 and 9.
In large specimens of P. speciosa from USA and Japan (ZIHU-1920, 1921) the ligula is shallow and the nuchal papilla invisible. In the syntypes, which are small, whole mounted specimens, both ligula and nuchal papilla may be present, although not very distinct and the observations are admittedly somewhat uncertain. We here tentatively treat the syntypes and the other specimens as conspecific; collection and examination of topotypes of various sizes would clearly be of great value .
Two Japanese specimens (ZIHU-1920, 1921) are herein referred to P. speciosa , and share, e.g. the poorly defined nuchal papilla, the undivided proboscis with lateral rows of large papillae, the short dorsal cirrophores, and the presence of large single main tooth on the rostrum of the chaetal shaft. Nevertheless, common Pacific and Atlantic distributions is unusual, and examination of further specimens from a large range of localities are needed.
PARANAITIS USCHAKOVI EIBYE- JACOBSEN, 1991
( FIGS 9 View Figure 9 , 34 View Figure 34 , 35 View Figure 35 , 37 View Figure 37 )
Paranaitis uschakovi Eibye-Jacobsen, 1991: 129 View in CoL . (replacement name for Paranaitis caecum Uschakov, 1972 View in CoL .
Paranaitis caecum Uschakov, 1972: 142–143 View in CoL , pl. 7, figs 5–7. Junior homonym to Paranaitis caeca ( Moore, 1903) View in CoL .
Material examined
Holotype ( ZIR 16140), east of Honshu , 38∞38¢N, 141∞53¢E, 239 m; 1 specimen (ZIHU-1922), Otsuchi Bay , Iwate Honshu, Japan, 39∞ 20.9 N 141∞58.5¢E, shell sand, 56 m, 25 May 1998, coll. T.K.; 1 specimen (ZIHU- 1923), Otsuchi Bay, 39∞20.5¢N, 141∞57.4¢E, sandy mud, 45 m, 25 May 1998, coll. T.K.
Description
Holotype, largest known specimen, 20 mm long, 2 mm wide, for 62 segments ( Uschakov, 1972); see Figure 9 View Figure 9 for measurements of other specimens. Live animals white, with single large rounded brown spots on inner parts of dorsal cirri, forming two longitudinal dorsal lines. Pigmentation disappears following preservation. Body short, dorso-ventrally flattened, posteriorly tapered. Prostomium rounded triangular, with shallow ligula ( Fig. 34A View Figure 34 ). Paired antennae and palps narrow, anteriorly directed. Eyes absent. Nuchal papilla small, rounded ( Fig. 34B View Figure 34 ). Nuchal organs rounded, rather large, situated ventrally, not dorsally visible. Proboscis lacking distinct papillae (examined by dissection; ZIHU-1923). Terminal ring with large number of poorly defined papillae. Inside of proboscis covered by small rounded, flat papillae. Paired large papillae present laterally inside ring.
Segments 1 and 2 indistinctly separated dorsally and ventrally. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered. Cirri of segment 1 reaching c. segment 5. Dorsal cirri of segments 2 and 3 reaching c. segment 8. Ventral cirri of segment 2 reaching c. segment 6. Segment 2 with ventral aciculae and c. six chaetae arising from small neuropodial lobes fused to ventral cirrophores. Segment 3 with neuropodial lobes with ventral aciculae and c. seven chaetae, with ventral cirri. Ventral aciculae present from segment 2. Dorsal aciculae of segment 2 and 3 absent.
Dorsal cirri of median segments kidney-shaped, symmetrical, wider than long ( Fig. 35A–D View Figure 35 ). Dorsal cirrophores dorsally prolonged, with weakly developed ciliary bands. Neuropodial lobes long, with supra-acicular lobes longer than subacicular lobes, with c. 25 chaetae. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, with single large main tooth on anterior side ( Fig. 37J View Figure 37 ). Ventral cirri oval, slightly pointed, about as long as neuropodia in median segments, slightly longer in posterior segments. Pygidial cirri short, oval ( Fig. 34C View Figure 34 ). Pygidial papilla present.
Habitat
Sandy bottom, 45–598 m.
Distribution
Known from east of Honshu, Japan.
Remarks
Paranaitis uschakovi View in CoL was originally described as Paranaitis caecum View in CoL by Uschakov (1972). In his revision of Eumida, Eibye-Jacobsen (1991) View in CoL transferred Eumida caeca, Moore, 1903 View in CoL ; to Paranaitis View in CoL . Since Paranaitis caecum Uschakov, 1972 View in CoL thereby became a junior homonym of Paranaitis caecum ( Moore, 1903) View in CoL , Eibye-Jacobsen introduced the new name Paranaitis uschakovi View in CoL for Uschakov’s species.
The examined specimens conform to P. uschakovi View in CoL and are characterized by the following unique combination of characters: absence of eyes, shallow but distinct ligula, neuropodial lobes with chaetae on segment 2, and white body with dark spots on inner part of dorsal cirri.
There has been some confusion regarding differences between the present species and P. caeca . We suggest that they may be separated by a number of morphological differences; see remarks for P. caeca for further information.
PARANAITIS INCERTAE View in CoL SEDIS PARANAITIS CAPENSIS ( DAY, 1960)
( FIG. 36 View Figure 36 )
Phyllodoce (Anaitis) capensis Day, 1960: 298 , fig. 5a–c. Phyllodoce (Paranaites) capensis (sic): Day, 1967: 148,
fig. 5.2q–s.
Paranaitis capensis: Pleijel, 1991: 258 .
Material examined
Holotype ( SAM-A20265 ), False Bay , South Africa, 34∞09.6¢S, 18∞29.3¢E, 31 m.
Description
Holotype complete specimen, 33 mm long, 1.8 mm wide, for 105 segments. Body colour of live specimens near posterior margin of prostomium. Nuchal papilla short, rounded, posteriorly situated in ligula. Nuchal organs not observed. Proboscis (probably dissected by Day) in poor condition, with distinct division separating proximal and distal part; proximal part covered with wide oval chitinous papillae, distal part with 5–6 rows of rounded tubercles. Terminal ring indistinct.
Segments 1 and 2 fused. Cirri of segment 1, dorsal and ventral cirri of segment 2, and dorsal cirri of segment 3 cylindrical, long and tapered. Cirri of segment 1 reaching c. segment 5. Dorsal cirri of segments 2 and 3 reaching c. segment 7. Ventral cirri of segment 2 reaching c. segment 5. Segment 2 without neuropodial lobes and chaetae. Segment 3 with small neuropodial lobes with c. six chaetae, with ventral cirri of similar shape and size as following segments. Aciculae of segments 2 and 3 unknown.
Dorsal cirri of median segments oval, longer than wide. Dorsal cirrophores short, distinct, symmetrical. Neuropodium with supra-acicular lobes longer than subacicular lobes, with 15–20 chaetae. Dorsal and ventral chaetae similar within single fascicle. Rostrum of chaetal shaft asymmetrical, single main tooth on anterior side, smaller tooth on posterior side. Ventral cirri with rounded ends, as long as neuropodial lobes. Pygidial cirri cylindrical with rounded ends, c. three times as long as wide ( Fig. 36B View Figure 36 ). Pygidial papilla present.
Habitat
Sand, shell and rocks, 55 m.
Distribution
Known only from False Bay and Cape ( Day, 1967), South Africa.
unknown. Preserved specimen uniformly creamy white. Body dorso-ventrally flattened, posteriorly tapered. Prostomium anteriorly rounded, c. 1.5 times as wide as long, with deep ligula ( Fig. 36A View Figure 36 ). Paired antennae and palps conical, laterally to posteriorly directed. Eyes medium-sized, rounded, with lenses, situated Remarks
Following Day, 1960, P. capensis differs from P. kosteriensis in the following characters: (1) more developed collar, (2) different proboscis, (3) longer dorsal cirri, and (4) different morphology of chaetal shafts. However, we cannot confirm any of these differences from examination of the holotype. In view of the poor condition of this specimen, of the lack of additional material, and of the distribution of the taxa, we avoid to synonymize P. capensis with P. kosteriensis . Further investigation based on newly collected South African specimens are required.
Annenkova NP. 1937. The Polychaeta Fauna of the northern part of the Japan Sea. Issledovaniya Morei SSSR 23: 139 - 216 (in Russian).
Berkeley E. 1924. Polychaetous annelids from the Nanaimo district. Part 2 Phyllodocidae to Nereidae. Contributions to Canadian Biology, Ottawa 2: 287 - 293.
Blake JA. 1994. 4. Family Phyllodocidae Savigny, 1818. In: Blake JA, Hilbig B, eds. Taxonomic atlas of the benthic fauna of the Santa Maria basin and western Santa Barbara channel, Vol. 4 - the Annelida, part 1, Oligochaeta and Polychaeta Phyllodocida (Phyllodocidae to Paralacydoniidae). Santa Barbara, CA: Santa Barbara Museum of Natural History, 115 - 186.
Day JH. 1960. The polychaete fauna of South Africa. Part 5. Errant species dredged off Cape Coasts. Annals of the South African Museum 45: 261 - 373.
Day JH. 1967. A monograph on the Polychaeta of Southern Africa. Part I, Errantia. London: Trustees of the British Museum (Natural History).
Day JH. 1973. New Polychaeta from Beaufort, with a key to all species recorded from North Carolina. NOAA Technical Report NMFS 375: 1 - 140.
Eibye-Jacobsen D. 1991. A revision of Eumida Malmgren, 1865 (Polychaeta: Phyllodocidae). Steenstrupia 17: 81 - 140.
Fauchald K. 1972. Benthic polychaetous annelids from deep water off western Mexico and adjacent areas in the eastern Pacific Ocean. Allan Hancock Monographs in Marine Biology 7: 1 - 575.
Gathof JM. 1984. Family Phyllodocidae Williams, 1851. In: Uebelacker JM, Johnson PG, eds. Taxonomic guide to the polychaetes of the northern Gulf of Mexico. Mobile, Alabama: Barry A. Vittor & Associates Inc., Chap. 19, 19 - 11, 19 - 42.
Hartman O. 1936. A review of the Phyllodocidae (Annelida Polychaeta) of the coast of California, with descriptions of nine new species. University of California Publications in Zoology 41: 117 - 132.
Hartman O, Reish DJ. 1950. The marine annelids of Oregon. Oregon State Monographs, Studies in Zoology 6: 1 - 64.
Hartman O. 1959. Catalogue of the polychaetous annelids of the world. Part I. Allan Hancock Foundation Publications, Occasional Paper 23: 1 - 353.
Hartman O. 1968. Atlas of the errantiate polychaetous annelids of California. Los Angeles, CA: Allan Hancock Foundation, University of Southern California Press.
Moore JP. 1903. Polychaeta from the coastal slope of Japan and from Kamchatka and Bering Sea. Proceedings of the Academy of Natural Sciences, Philadelphia 55: 401 - 490.
Moore JP. 1909. The polychaetous annelids dredged by the U. S. S. ' Albatross' off the coast of southern California in 1904. I. Syllidae, Shaerodoridae, Hesionidae and Phyllodocidae. Proceedings of the Academy of Natural Sciences of Philadelphia 61: 321 - 351.
Pettibone MH. 1963. Marine polychaete worms of the New England region. Bulletin of the United States National Museum 227: 1 - 356.
Uschakov PV. 1972. Polychaeta 1. Polychaetes of the suborder Phyllodociforma of the Polar Basin and the north-western part of the Pacific. Fauna SSSR. Zoologicheskii Institut Akademii Nauk SSSR, Moskva and Leningrad 102: 1 - 271 (in Russian, translated by the Israel Program for Scientific Translation, Jerusalem, 1974).
Webster HE. 1879. Annelida chaetopoda of the Virginian coast. Transactions of the Albany Institute 9: 202 - 269.
Webster HE, Benedict JE. 1884. The Annelida Chaetopoda from Provincetown and Wellfleet, Mass. Report of the United States Commissioner of Fisheries 1884: 699 - 747.
Webster HE, Benedict JE. 1887. The Annelida Chaetopoda, from Eastport, Maine. Report of the United States Commissioner of Fisheries 1885: 707 - 755.
Zachs I. 1933. Polychaeta of the North Japanese Sea. Issledovaniya Morei SSSR 19: 125 - 137.
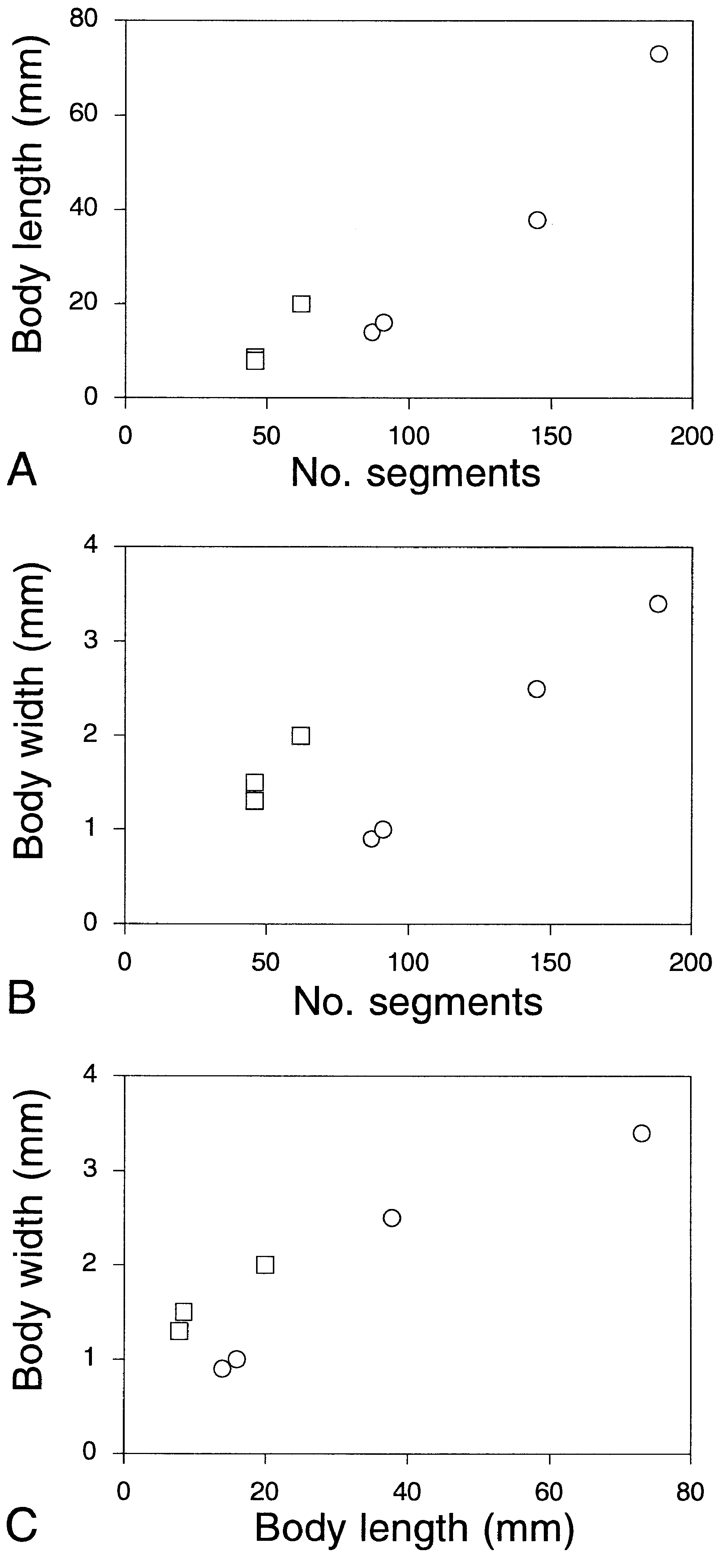
Figure 9. Paranaitis caeca and P. uschakovi, specimens from Japan. Relationships between A, number of segments and body length, B, number of segments and body width, and C, body length and body width. Circles represent P. caeca, and squares P. uschakovi.
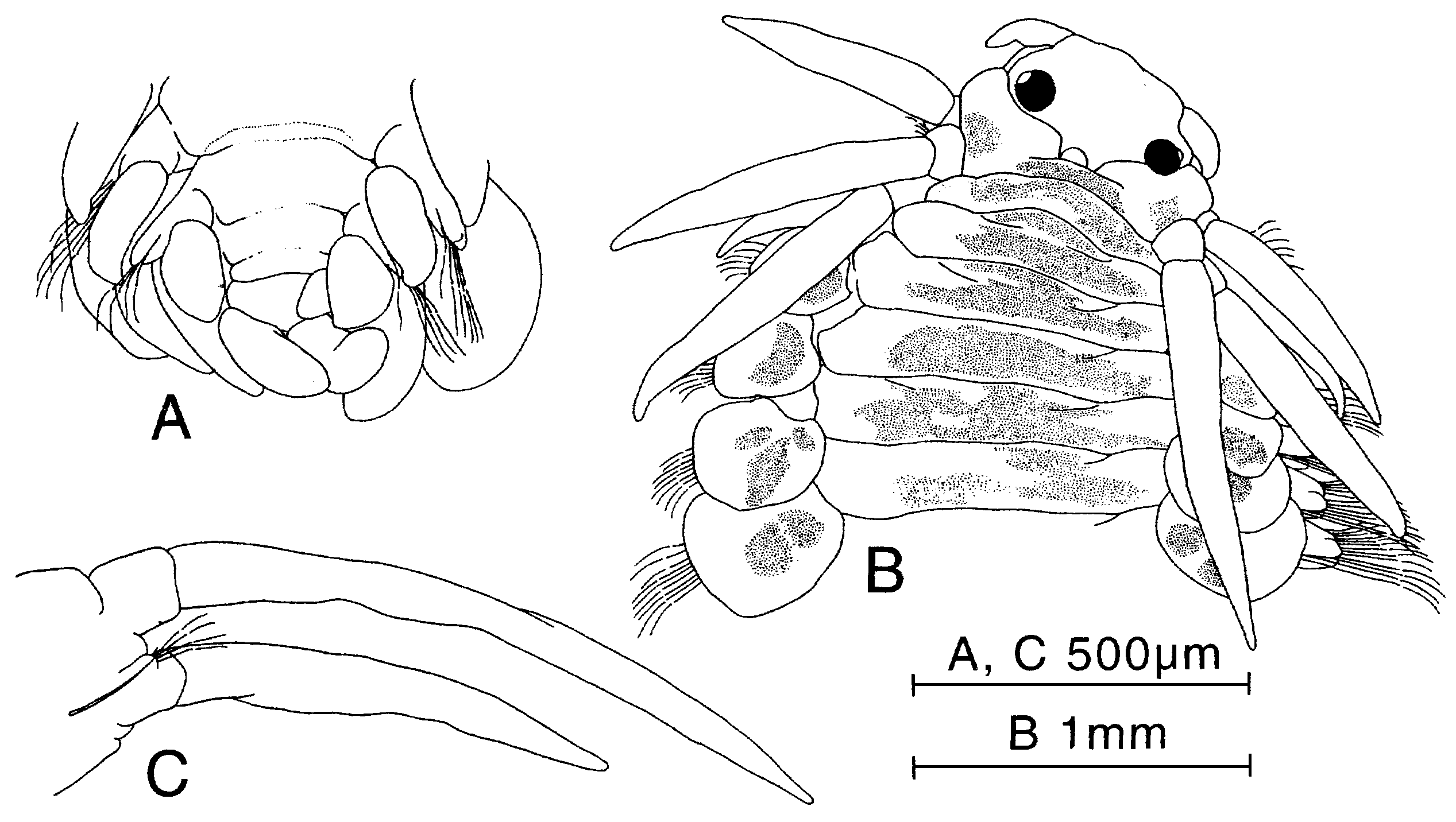
Figure 23. Paranaitis moritai sp. nov. A, holotype (ZIHU-1925). B, paratype (ZIHU-2291). A, posterior end, ventral view. B, anterior end, dorsal view. C, parapodium of segment 2, anterior view.
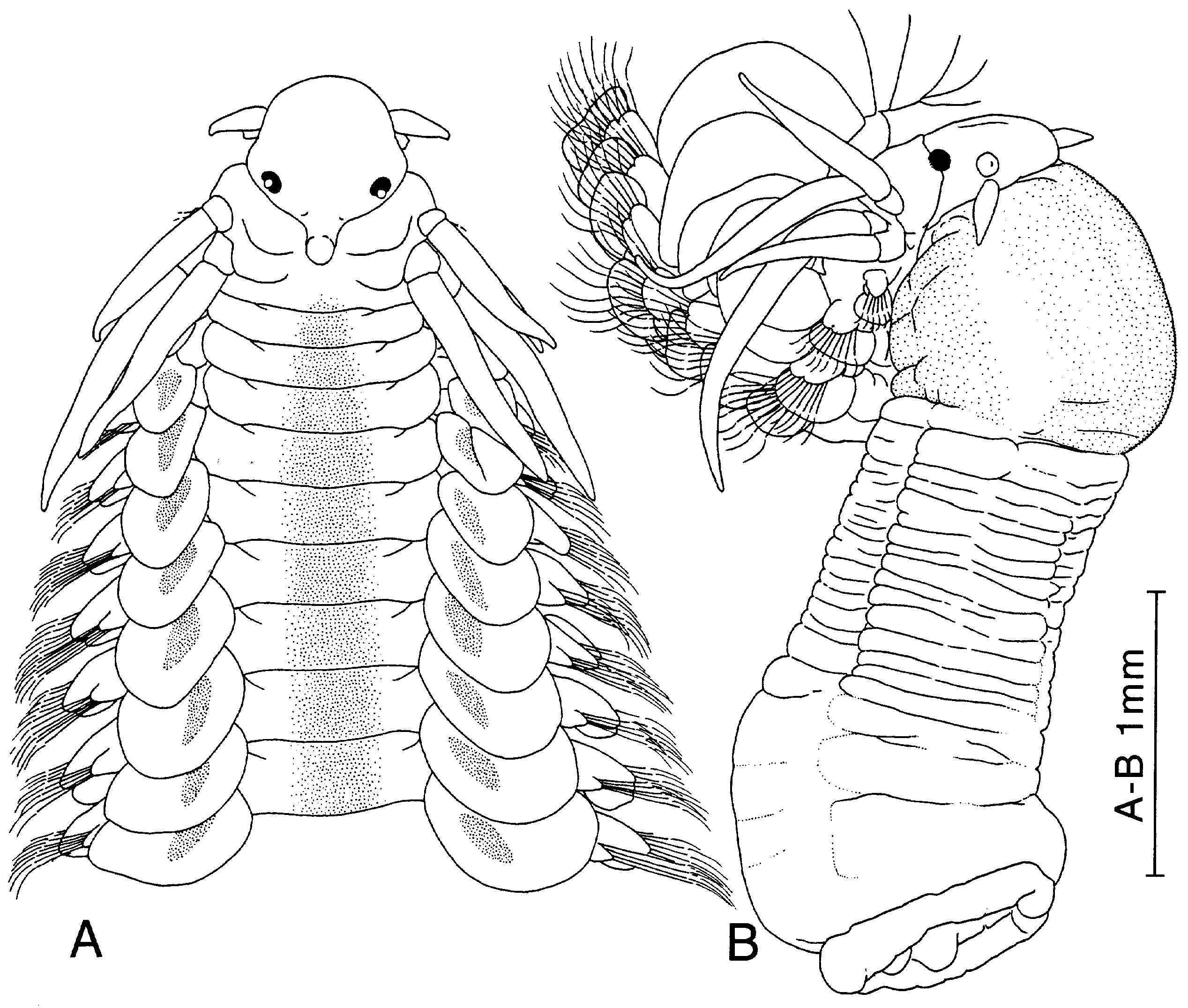
Figure 24. Paranaitis polynoides. A, specimen from Washington, USA (USNM-32461). B, specimen from Santa Monica Bay, California (ZIHU-2016). A, anterior end, dorsal view. B, anterior end with fully everted proboscis, lateral view.

Figure 25. Paranaitis polynoides, specimen from Santa Monica Bay, California (ZIHU-2016). A, cirri of segment 1, anterior view. B, parapodium of segment 2, anterior view. C, parapodium of segment 3, anterior view. D, parapodium of segment 4, anterior view. E, parapodium of segment 5, anterior view. F, parapodium of segment 21, anterior view. G, same, posterior view. H, posterior end of body, ventral view.

Figure 26. Paranaitis polynoides, specimens from USA. Relationships between A, number of segments and body length, B, number of segments and body width, and C, body length and body width.
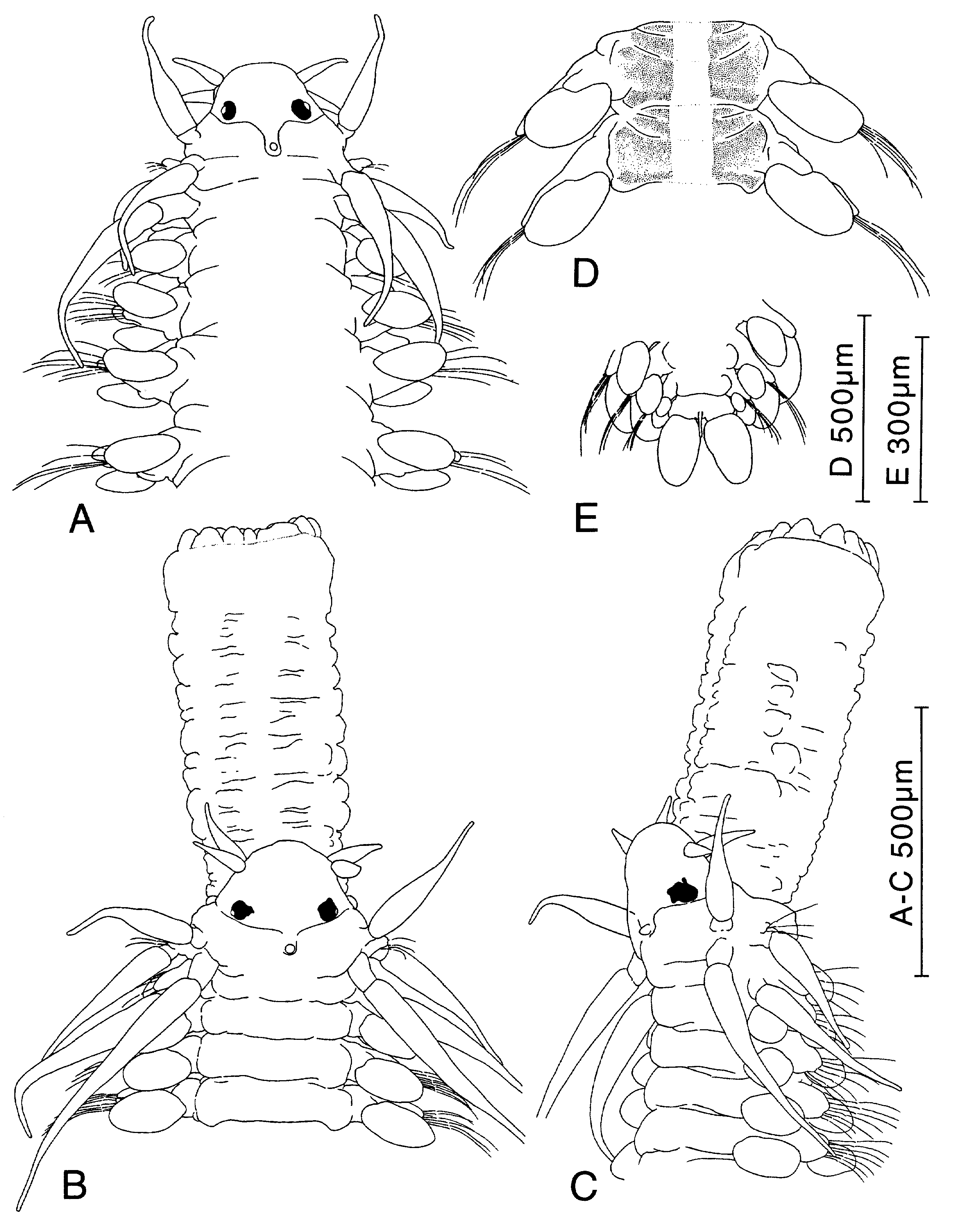
Figure 27. Paranaitis pumila sp. nov. A, D, E, holotype; B, C, specimen from Otsuchi Bay, Japan (ZIHU-1974). A, anterior end, dorsal view. B, anterior end with fully everted proboscis. C, same, lateral view. D, median segments, dorsal view. E, posterior end, ventral view.
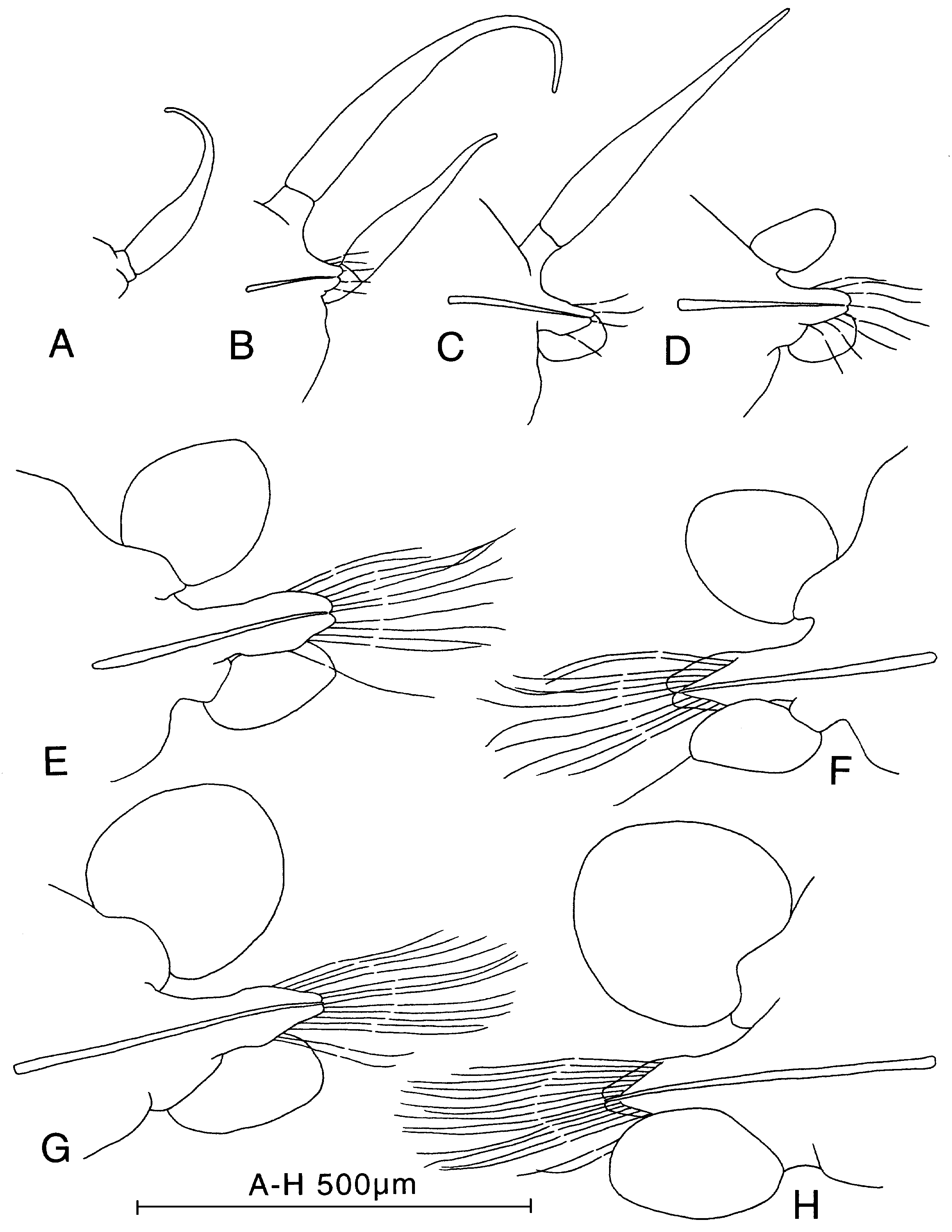
Figure 28. Paranaitis pumila sp. nov., paratype (ZIHU-1379). A, cirri of segment 1, anterior view. B, parapodium of segment 2, anterior view. C, parapodium of segment 3, anterior view. D, parapodium of segment 4, anterior view. E, parapodium of segment 17, anterior view. F, same, posterior view. G, parapodium of segment 44, anterior view. F, same, posterior view.

Figure 29. Paranaitis pumila sp. nov., type series. Relationships between A, number of segments and body length, B, number of segments and body width, and C, body length and body width.
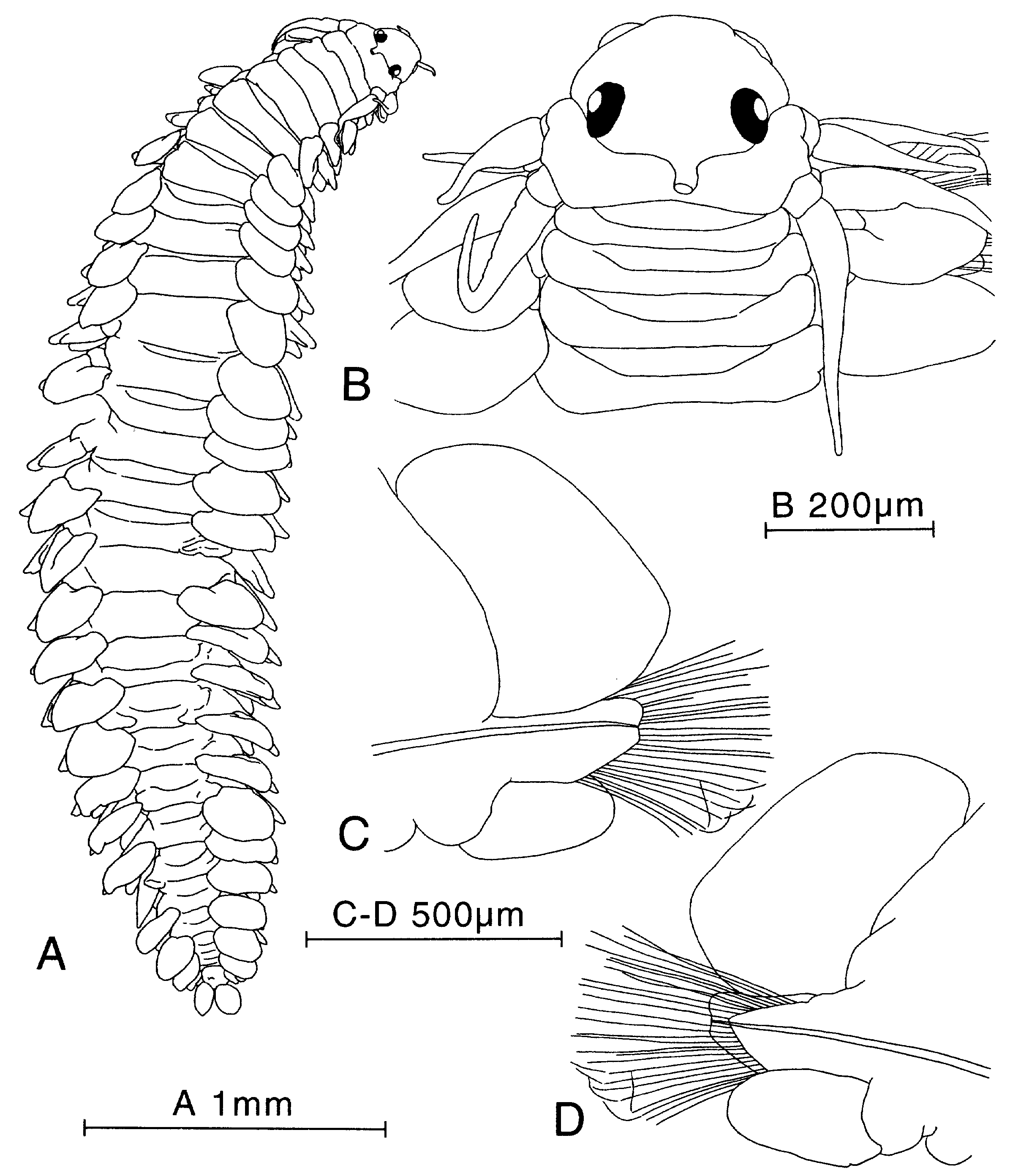
Figure 30. Paranaitis speciosa, syntype (USNM-380). A, entire animal, dorsal view. B, anterior end, dorsal view. C, parapodium of median segment, anterior view. D, same, posterior view.
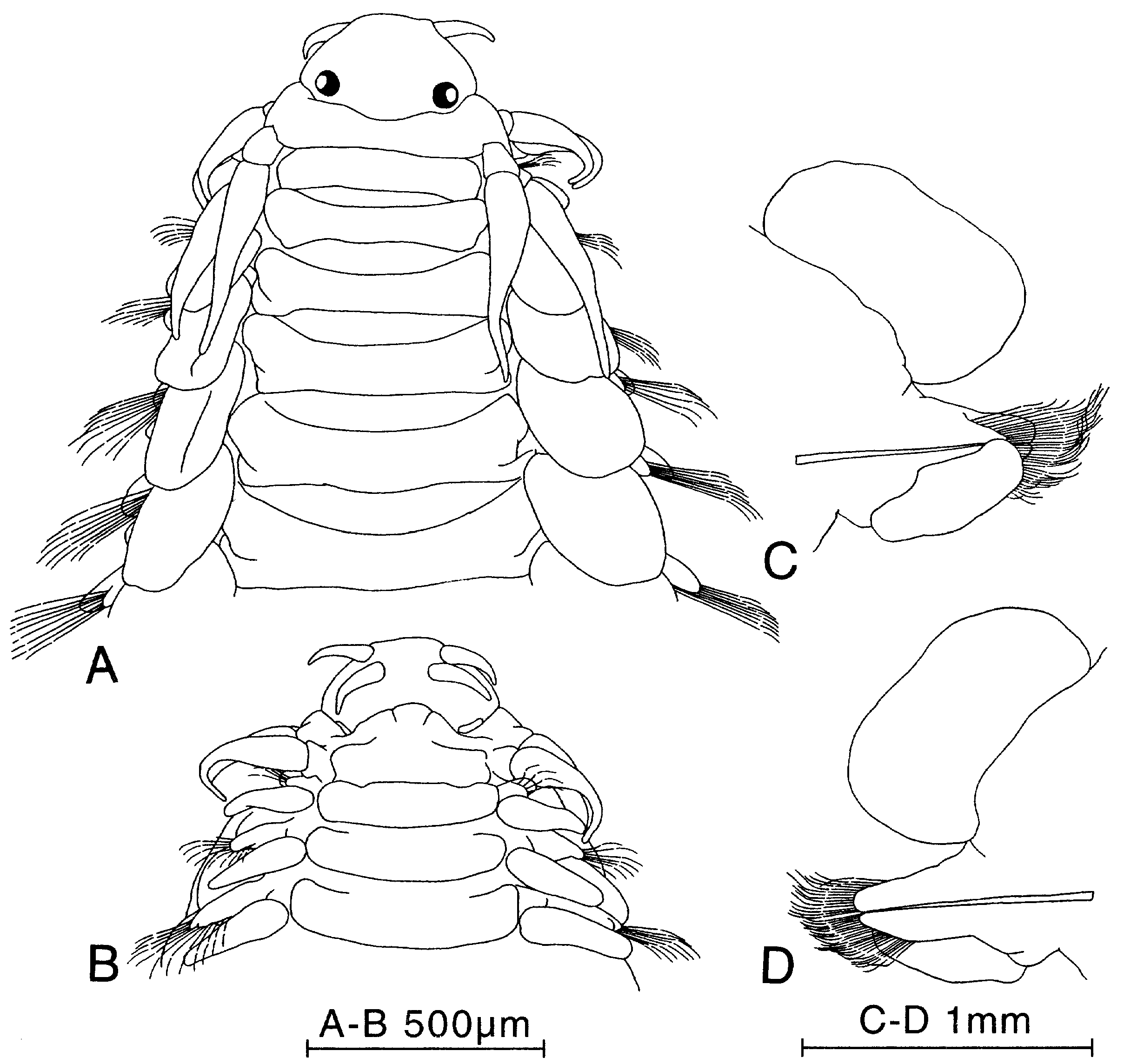
Figure 31. Paranaitis speciosa, specimen from North Carolina, USA (Mary Petersen’s personal collection). A, anterior end, dorsal view. B, same, ventral view. C, parapodium of segment 18, posterior view. D, same, anterior view.
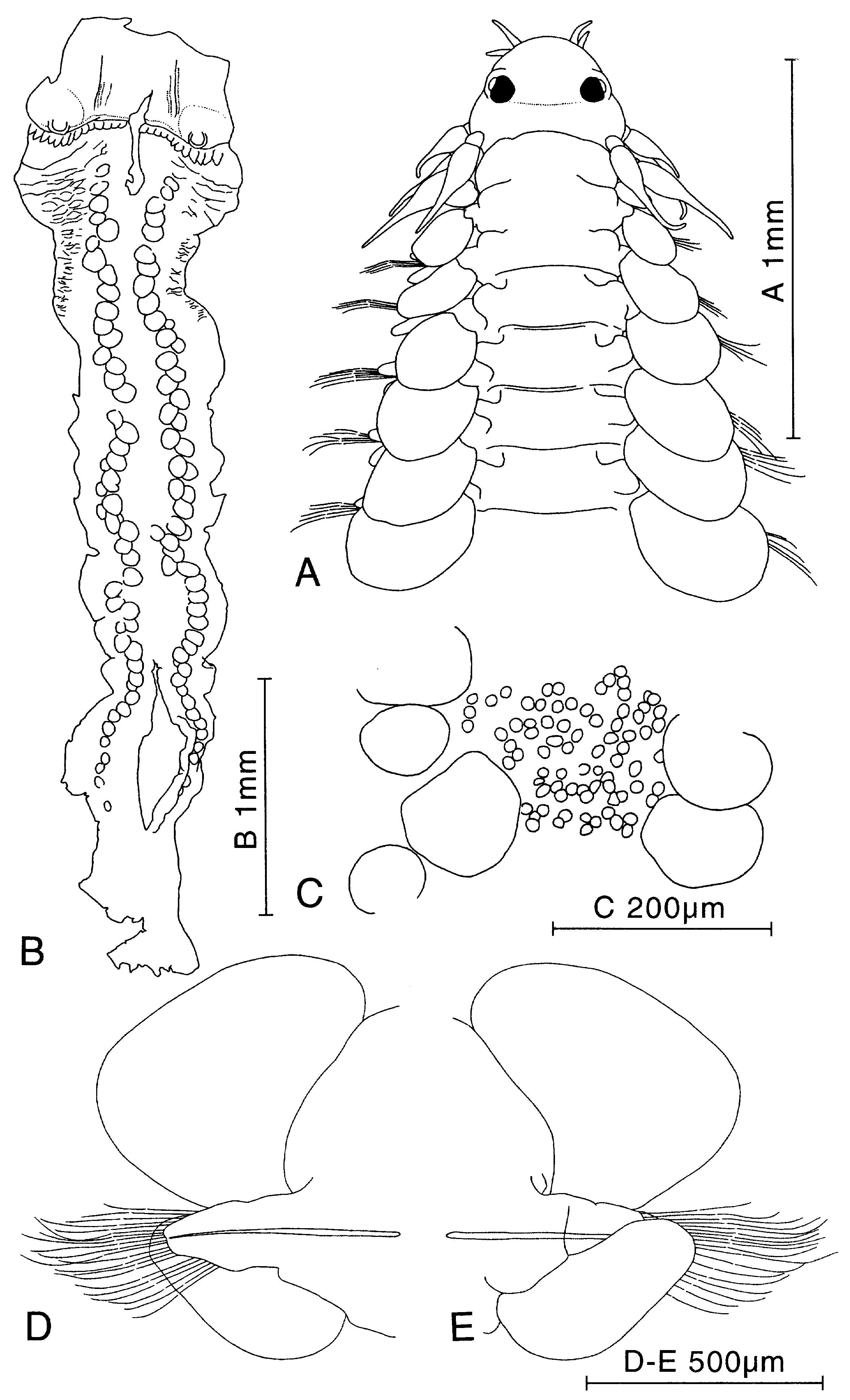
Figure 32. Paranaitis speciosa. Specimen from Japan (ZIHU-1921). A, anterior end, dorsal view. B, dissected proboscis, ventral view (median area corresponds to dorsal part). C, proboscis papillae. D, parapodium of segment 32, anterior view. E, same, posterior view.

Figure 33. Paranaitis speciosa, specimens from USA and Japan. Relationships between A, number of segments and body length, B, number of segments and body width, and C, body length and body width. Circles represent specimens from USA, squares, Japan.

Figure 34. Paranaitis uschakovi, specimen from Otsuchi Bay, Japan (ZIHU-1923). A, anterior end, dorsal view. B, same, lateral view. C, posterior end, ventral view.

Figure 35. Paranaitis uschakovi, specimen from Otsuchi Bay, Japan (ZIHU-1923). A, parapodium of segment 22, anterior view. B, same, posterior view. C, parapodium of segment 32, anterior view. D, same, posterior view.

Figure 36. Paranaitis capensis, holotype. A, anterior end, dorsal view. B, posterior end, ventral view.
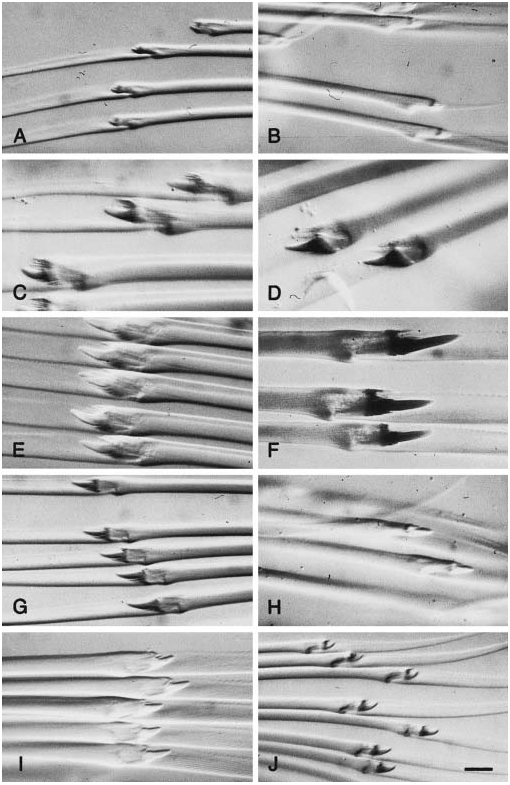
Figure 37. Chaetae of species of Paranaitis. A, P. wahlbergi, specimen from Sweden, ZIHU-2014. B, P. caeca, specimen from Japan, ZIHU-1919. C, P. kosteriensis, specimen from Sweden, SMNH-22547, dorsal chaetae. D, P. kosteriensis, same specimen, ventral chaetae. E, P. misakiensis sp. nov., paratype, ZIHU-1381. F, P. polynoides, ZIHU-2016. G, P. pumila sp. nov., paratype, ZIHU-1379. H, P. speciosa, syntype USNM 380. I, P. speciosa, specimen from Japan, ZIHU-1921. J, P. uschakovi, specimen from Japan, ZIHU-1923.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Paranaitis
| Kato, Tetsuya & Pleijel, Fredrik 2003 |
Paranaitis uschakovi
| Eibye-Jacobsen D 1991: 129 |
Paranaitis polynoides: Fauchald, 1972: 52
| Gathof JM 1984: 19 |
| Fauchald K 1972: 52 |
Paranaitis caecum
| Uschakov PV 1972: 143 |
Phyllodoce (Anaitis) capensis
| Day JH 1967: 148 |
| Day JH 1960: 298 |
Paranaitis speciosa: Hartman, 1959: 158
| Gathof JM 1984: 19 |
| Day JH 1973: 22 |
| Pettibone MH 1963: 75 |
| Hartman O 1959: 158 |
Phyllodoce (Anaitis) polynoides:
| Annenkova NP 1937: 156 |
Paranaitis polynoides:
| Blake JA 1994: 164 |
| Uschakov PV 1972: 1141 |
| Hartman O 1968: 291 |
| Hartman O 1959: 158 |
| Hartman O 1936: 117 |
Anaitis polynoides
| Zachs I 1933: 127 |
| Berkeley E 1924: 287 |
| Moore JP 1909: 342 |
Anaitis speciosa
| Webster HE & Benedict JE 1887: 710 |
| Webster HE & Benedict JE 1884: 702 |
| Webster HE 1879: 132 |