
Sabellaria isumiensis, Nishi, Eijiroh, Bailey-Brock, Julie Helen, Santos, Andre Souza Dos, Tachikawa, Hiroyuki & Kupriyanova, Elena K., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199292 |
|
DOI |
https://doi.org/10.5281/zenodo.5620675 |
|
persistent identifier |
https://treatment.plazi.org/id/707B8788-FFC8-FFCE-FF75-0A6C7B29FA91 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellaria isumiensis |
| status |
sp. nov. |
Sabellaria isumiensis View in CoL n. sp.
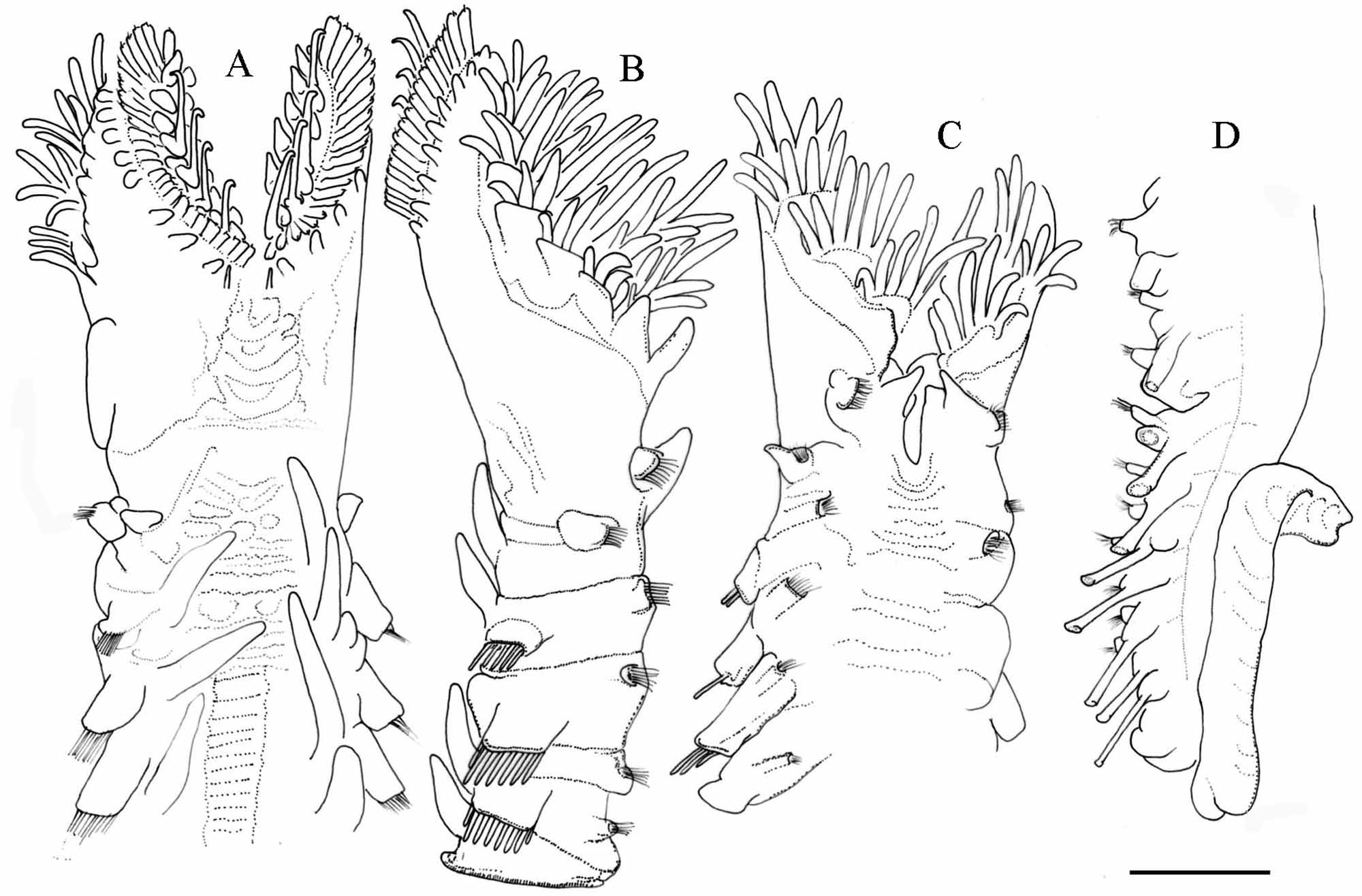
Figs. 10 View FIGURE 10 (A–E), 11 (A–D), 12 (A–G), 13 (A–F).
Sabellaria View in CoL cf. chandraae— Nishi & Kato, 2002: 12 –14 (non de Silva, 1961)
Material examined. Holotype CMNH-ZW 0 1630, collected at Kohazuki beach, Iwawata, Onjuku, Boso Peninsula, intertidal zone of rocky shore, 14 June 2003, by hand, coll. J. Takayama. Paratypes, 20 specimens (AM W.36825; BPBM-R3417; CBM-ZW-1026, 1027; CMNH-ZW 01629; CYPY-POLY 1030, 1031, 1032; NHM 2010.221, 222; MBM 179998, 179999; SAMA E3730; SMF 19450; USNM 1143566; ZIHU 3899, 3900; ZMUC-POL-2114), collection data same as for holotype.
Description. Holotype, immature complete specimen, 20.0 mm long, 1.5 mm wide at widest part of opercular crown, 2.0 mm wide at parathoracic chaetigers. Anterior end of opercular stalk completely divided into 2 bilaterally symmetrical lobes ( Fig. 11 View FIGURE 11 A, C) narrowing anteriorly, sloping posteriorly towards dorsal midline ( Fig. 11 View FIGURE 11 A, B). Operculum dish-shaped, flat to slightly concave.
Paleae bright yellow, with light pink or colourless bases (embedded in tissue). Opercular paleae in three rows (outer, middle, inner) on each lobe (Fig. 13 A). Outer paleae ca. 0.1 mm long and ca. 0.05 mm wide, curved slightly outward from longitudinal axis of shaft (Fig. 13 B), inner surface of their blade with ca. 50 fine closely-spaced thecal bands ( Fig. 12 View FIGURE 12 C). Distal ends of outer paleae with a median pilose plume and 2–3 pairs of lateral distal teeth with sharp tips (N=15, average=2.67, SD=0.62: paratype CMNH ZW 01629; Fig. 12 View FIGURE 12 A– C). Plume short, pilose (i.e., denticulate with 2–4 small lateral teeth on each side), emerging from top or middle of central tooth (N=15, average=3.0, SD=0.53: paratype CMNH ZW 01629), decreasing in size distally ( Fig. 12 View FIGURE 12 A–C). Middle paleae of two types, long and short. Long arranged in five pairs, with erect blades tapering slowly to blunt tips recurved towards outer margin of crown ( Figs. 12 View FIGURE 12 D, 13 C). Short arranged in five to six pairs, geniculate, with thick wide blades terminating distally in blunt tips ( Fig. 12 View FIGURE 12 E). Blades of long and short kinds of paleae with transverse thecal bands. Inner paleae in 9 or 10 pairs, similar in shape to the short form of middle paleae, but usually smaller in size with shorter blades ( Fig. 12 View FIGURE 12 F). Surface of excavated blades of inner palea with transverse thecal bands ( Fig. 12 View FIGURE 12 F); blades semi-transparent, colorless, slender, straight, with blunt tips. One to three dorsal nuchal spines arising from each side of dorsal midline ( Figs. 11 View FIGURE 11 A, 13 B).
Anterior outer margin of opercular crown with 9–10 fleshy conical papillae in single row on each side ( Fig. 11 View FIGURE 11 A, B). Ventral margin of buccal cavity with 10–15 longitudinal rows of feeding tentacles on each side ( Fig. 11 View FIGURE 11 B, C). A pair of grooved tentacles located after the stoma. Building organ U-shaped, with conical cirri, forming upper lip on ventral side ( Fig. 11 View FIGURE 11 C). Inferior lobes of second chaetiger with fascicle of ca. 8 bipinnate capillary chaetae and conical lateral cirrus. Superior lobes with achaetous conical cirri along lateral margin of opercular lobe, with dorsal conical branchia ( Fig. 11 View FIGURE 11 A, B).
Three parathoracic chaetigers with 8–10 stout oar-shaped chaetae (Fig. 13 D) and spine-like companion chaetae in 2 or 3 rows on each neuropodium and ca. 10 chaetae on each notopodium. Abdominal region with 25 chaetigers with bundles of 10–15 long simple capillaries and finely serrated neurochaetae (Fig. 13 E), and elongate conical ventral cirri with an uncinigerous torus. Neurochaetal conical cirri arising from base of bundle in first abdominal segment. In the first three abdominal chaetigers the conical cirri have lateral projection, after third abdominal chaetiger, the cirri decrease to short lobe. Uncinigerous tori with chaetae in single row. Notopodial uncini bi-pectinate with 6 or 7 rows of teeth superimposed (Fig. 13 F). Twelve anterior-most abdominal chaetigers with dorsal branchiae, diminishing in size posteriorly ( Fig. 11 View FIGURE 11 D). The branchiae short in anterior chaetigers, by third parathoracic the branchiae become longer, and maintain the length until third abdominal chaetiger. After third chaetiger the branchiae decrease towards posterior chaetigers until they disappear in last abdominal chaetigers. Caudal region septate composed of fused achaetous segments, 3.0 mm long and bent back on ventral with anus and 25–30 anal appendices.
Eyespots brownish to reddish in color on each filament of compound feeding tentacles. Brown eyespots also on opercular peduncle, on inner surface of serial conical palps extended to mid-ventral indentation, on prostomium, on peristomium between compound feeding tentacles, on median cirrus (from base to upper edge), on thoracic region, and on parathoracic region (a large patch of numerous eyespots in ventral side of third chaetiger). Black eyespots on ventral side of abdomen and in caudal region. These eyespots absent on the first abdominal chaetiger, after the second abdominal chaetiger they form C-shaped pads, after 5th abdominal chaetiger the eyespots increase in number and become organized in patches of numerous eyespots.
Variation. Paratypes with four or six pairs of the long form of middle row paleae and five or six pairs of the short form of middle row paleae. Inner surface of the short form depressed in basal part ( Fig. 12 View FIGURE 12 D). The paleae in paratypes 15.0–30.0 mm long and 0.8–2.2 mm wide at parathoracic chaetigers. Abdominal region with 10–30 chaetigers. Anterior-most 10–15 abdominal chaetigers with dorsal branchiae decreasing in size posteriorly. The cauda 3.0–5.0 mm in length.
Tube. Tubes constructed of sand and shell debris, with inner side coated with a thin membrane. Tubes highly gregarious, forming large honey-comb colonies over 2.0 m wide. Tube orifice 1.0–1.5 mm in diameter.
Remarks. Among 35 species of Sabellaria described worldwide (including S. isumiensis n. sp.), 21 are reported to have both nuchal spines and long middle paleae (Table 1). However, to confirm the presence of nuchal spines, observation of sectioned specimens is required, because nuchal spines can be very small and inserted in the opercular stalk. Sabellaria isumiensis n. sp. differs from all other members of the genus by having both straight nuchal spines and long middle paleae with distal tips curved towards the periphery of the crown (Table 1). The new species resembles closely S. chandraae (de Silva, 1961) from Sri Lanka and S. ranjhi ( Hasan, 1960) from Pakistan, in having long and short middle paleae and denticulate outer paleae (Figs.
13, 14). Sabellaria isumiensis n. sp. can be distinguished from those two species by the outer paleae morphology, i.e., pilose median tooth of S. chandraae has 5–8 lateral teeth ( Fig. 6 View FIGURE 6 A, E), but the medial tooth of S. isumiensis n. sp. has 2, 3 or 4 lateral teeth ( Fig. 12 View FIGURE 12 A–C). These three species are easily distinguished by the shape of the short middle palea: S. chandraae and S. ranjhi have clavate short middle paleae, while S. isumiensis n. sp. have elliptical ones.
Etymology. The species epithet S. isumiensis is chosen for the type locality, Isumi area, Onjuku and Katuura, Boso Peninsula, Chiba Prefecture.
Type locality. Onjuku to Katsuura, Isumi area, Pacific side of Boso Peninsula, Honshu, Japan.
FIGURE 13. Sabellaria isumiensis n. sp. Scanning electron micrographs. Opercular crown (A), nuchal spine and outer paleae (B), middle palea (C), 3rd parathoracic oar-shaped chaetae (D), abdominal chaetae (E), and posterior abdominal uncini (F).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sabellaria isumiensis
| Nishi, Eijiroh, Bailey-Brock, Julie Helen, Santos, Andre Souza Dos, Tachikawa, Hiroyuki & Kupriyanova, Elena K. 2010 |
Sabellaria
| Nishi 2002: 12 |