
Apteronotus ferrarisi, Santana & Vari, 2013
|
publication ID |
https://doi.org/ 10.1111/zoj.12022 |
|
persistent identifier |
https://treatment.plazi.org/id/701A87DA-BD18-D06E-51F7-FEAF224DFCA5 |
|
treatment provided by |
Marcus |
|
scientific name |
Apteronotus ferrarisi |
| status |
sp. nov. |
APTERONOTUS FERRARISI SP. NOV.
( FIGS 5 View Figure 5 , 9 View Figure 9 , 10 View Figure 10 ; TABLE 1)
Diagnosis: Apteronotus ferrarisi is distinguished from A. anu by the condition of the fifth infraorbital (absent versus present as a tubular laterosensory canal segment), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of scales above the lateral line at the midbody (10–13 versus seven to nine), the number of scales along the lateral line (73–80 versus 59–63), the number of caudal-fin rays (15–17 versus 10–13), the extent of the middorsal groove (extending two to six scales beyond the vertical through the posterior terminus of the anal fin versus seven to eight) and the tail length (6.9–9.9% of LEA versus 17.1–37.5%). Apteronotus ferrarisi differs from A. baniwa in the number of scales above the lateral line at the midbody (10–13 versus seven to eight), the number of scales along the lateral line (73–80 versus 61–70), the mouth length (49.4–56.4% of HL versus 36.6–47.2%), the tail length (6.9-9.9% of LEA versus 13.3–24.2%), and the tail depth (22.2– 28.7% of CL versus 8.2–15.3%). Apteronotus ferrarisi is differentiated from A. galvisi by the orbital diameter (4.0–5.9% of HL versus 6.4–11.3%) and the tail length (6.9-9.9% of LEA versus 11.8–15.9%). Apteronotus ferrarisi is distinguished from A. leptorhynchus by the mouth length (49.4–56.3% of HL versus 35.1–48.8) and the tail length (6.9-9.9% of LEA versus 17.1–18.6%). Apteronotus ferrarisi differs from A. macrostomus in the tail length (6.9-9.9% of LEA versus 13.9–20.6%), the number of caudal-fin rays (15–17 versus 18–21), and the number of total vertebrae (61–71 versus 75). Apteronotus ferrarisi is differentiated from A. pemon by the mouth length (49.4– 56.3% of HL versus 30–44.5%), the tail length (6.9-9.9% of LEA versus 13.0–17.5%), the tail depth (22.2–28.7% of CL versus 9.9–15.4%), and the number of caudal-fin rays (15–17 versus 18–20). Apteronotus ferrarisi is distinguished from A. rostratus by the condition of the fifth infraorbital (absent versus present as a tubular laterosensory canal segment), the condition of the lateral ethmoid (ossified versus cartilaginous), the tail length (6.9-9.9% of LEA versus 10.6–12.4%), and the number of scales along the lateral line (73–80 versus 63–67). Apteronotus ferrarisi differs from A. spurrellii in the condition of the fifth infraorbital (absent versus present as a tubular laterosensory canal segment), the condition of the lateral ethmoid (ossified versus cartilaginous), the mouth length (49.4–56.3% of HL versus 42.9–48.4%), the tail length (6.9-9.9% of LEA versus 12.4–28.7%), the number of scales above the lateral line at the midbody (10–13 versus eight to nine), the number of anal-fin rays (150–167 versus 171–173), and the number of caudal-fin rays (15–17 versus 14).
Description: Head and body shape and pigmentation illustrated in Figures 9 View Figure 9 and 10 View Figure 10 . Morphometrics for holotype and paratypes presented in Table 1. Body laterally compressed, greatest body depth located at, or slightly posterior to, abdominal cavity. Dorsal profile of body nearly straight. First perforated scale of lateral line located above pectoral-fin origin. Lateral line extending posteriorly to base of caudal fin. Scales along lateral line 73(1), 74(1), 77*(2), or 80(1). Scales above lateral line to mid-dorsal line at midbody 10(2), 11*(2), or 13(1).
Head laterally compressed, widest at opercular region and deepest at nape. Dorsal profile of head nearly straight but with snout longer in mature males ( Fig. 9 View Figure 9 ) than mature females ( Fig. 10 View Figure 10 ). Eye small, located laterally on head and completely covered by thin membrane. Anterior naris located at end of small tube and close to tip of snout. Posterior naris ellipsoidal, without tube and positioned closer to tip of snout than to anterior margin of eye. Mouth terminal with rictus extending posterior of vertical through anterior border of eye. Branchial opening located anterior to vertical through pectoral-fin insertion. Anus and urogenital papillae located posterior to vertical through eye and not varying ontogenetically in position.
Pectoral fin elongate, with 13(1), 15*(1), 16(2), or 17(1) rays. Anal-fin origin located anterior to vertical through posterior margin of opercle. Unbranched anal-fin rays 21(1), 23(1), 24(1), 25*(1), or 33(1); total anal-fin rays 150(1), 153*(1), 156(1), or 167(1). Tail compressed and short; ending in small, elongate, caudal fin. Caudal-fin rays 15*(3) or 17(1).
Origin of dorsal sagittal electroreceptive filament located on posterior half of body. Filament inserted into narrow mid-dorsal groove and extending 2(3), 4*(1), or 6(1) scales posterior of vertical through posterior terminus of anal fin. Precaudal vertebrae 17(2) or 18*(3). Total vertebrae 61(1), 68*(1), 70(2), or 71(1).
Secondary sexual dimorphism: Sexually mature males (N = 2, 231– 274 mm TL; Fig. 9 View Figure 9 ) and females (N = 3, 94– 111 mm TL, Fig. 10 View Figure 10 ) of A. ferrarisi differ in diverse components of head morphology. The length of the snout, mouth length, and the distance from the posterior naris to the eye are proportionally longer in mature males than in mature females ( Table 1).
Coloration in alcohol: Body dark and head dark brown. Prominent pale, broad, longitudinal band extending from chin along dorsal midline of head and body to beginning of dorsal filament. Pectoral and anal-fin rays dark brown and inter-radial membranes translucent. Pale band encircling base of caudal fin. Caudal fin pale at base and dark brown posteriorly.
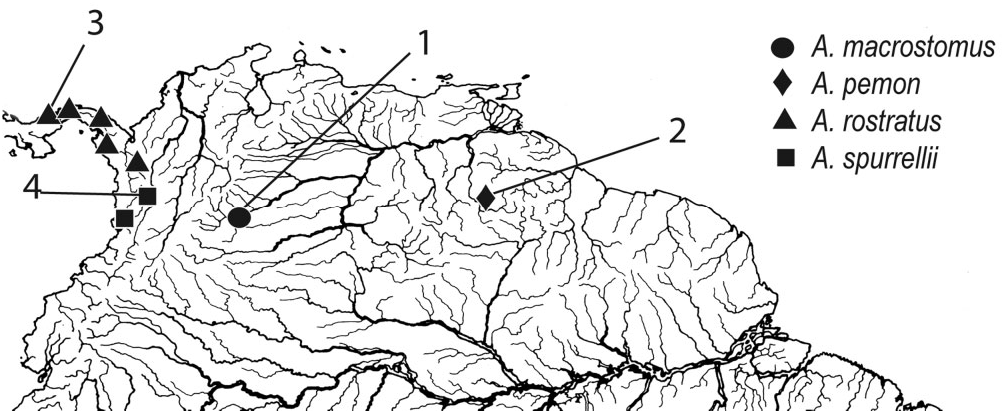
Distribution: Apteronotus ferrarisi is known from the type locality in the Río Yaracuy and also occurs in the adjoining Río Aroa, both of which are coastal Caribbean versant drainages of northern Venezuela ( Fig. 5 View Figure 5 ). These rivers are a centre of endemism within the Tocuyo-Yaracuy subprovince as defined by Rodríguez-Olarte et al. (2009: 87), with 24 endemic fish species previously reported in these basins.
Etymology: The species name, ferrarisi , is in honour of Carl J. Ferraris Jr, in recognition of his many contributions to our knowledge of tropical freshwater fishes worldwide and his invaluable assistance to the authors, particularly the second author, over the years.
Remarks: The report of Apteronotus sp. from the Tocuyo-Yaracuy region by Rodríguez-Olarte et al. (2009: 73) is presumably based on A. ferrarisi , which is the only member of the genus known to occur in those basins.
Material examined: Holotype: Venezuela. Yaracuy. MCNG 55635 View Materials , male, 231 mm; Río Yaracuy basin (Caribbean coast), Quebrada Guaquira on Hacienda Guaquira, south-east of El Peñon Reservoir , 10°17′40″N, 68°39′35″W, collected by D. Rodríguez, A. Amaro, H. Aguldelho, J. Coronel & D. Taphorn, 23.x.2005 GoogleMaps . Paratypes: Venezuela. Yaracuy. MCNG 54584 View Materials , 1 View Materials (male), 160 mm; collected at holotype locality, 21.x.2005. MCNG 54597 View Materials , 2 View Materials (females), 176– 196 mm; collected at holotype locality. MCNG 52077 View Materials , 1 View Materials (male), 274 mm; Río Carabobo, Río Aroa drainage (Caribbean coast), 10°30′08″N, 68°47′02″W, D. Taphorn et al., 15.i.2004 GoogleMaps .
APTERONOTUS GALVISI DE SANTANA , MALDONADO- OCAMPO & CRAMPTON
( FIGS 5 View Figure 5 , 11 View Figure 11 ; TABLE 2)
Apteronotus galvisi de Santana, Maldonado-Ocampo & Crampton, 2007: 118 View in CoL , fig. 1 [original description; type-locality Río Cusiana, piedmont of Cordillera Oriental, 305–424 m asl, upper Río Meta; secondary sexual dimorphism]. – Maldonado-Ocampo et al., 2008: 213 [ Colombia]. – Urbano-Bonilla et al., 2009: 162 [ Colombia, Departamento de Casanare, Ríos Pauto, Cravo Sur, Cusiana, Upía]. – Machado-Allison et al. 2010: 242 [Río Meta]. – de Santana & Cox-Fernandes, 2012: 284 [Río Orinoco basin; occurrence in streams; presence of secondary sexual dimorphism].
Diagnosis: Apteronotus galvisi is differentiated from A. anu by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of scales above the lateral line at the midbody (11–12 versus seven to nine), and the tail length (11.8–15.9% of LEA versus 17.1–37.5%). Apteronotus galvisi is distinguished from A. baniwa by the number of scales above the lateral line at the midbody (11–12 versus seven to eight). Apteronotus galvisi differs from A. ferrarisi in the orbital diameter (6.4–11.3% of HL versus 4.0–5.9%) and the tail length (11.8–15.9% of LEA versus 6.9-9.9%). Apteronotus galvisi is differentiated from A. leptorhynchus by the number of caudal-fin rays (10–16 versus 17–21), the length of the anal-fin base (69.4–81.1% of LEA versus 81.7–83.7%), and the tail length (11.8–15.9% of LEA versus 17.1–18.6%). Apteronotus galvisi is distinguished from A. macrostomus by the number of caudal-fin rays (10–16 versus 18–21). Apteronotus galvisi differs from A. pemon in the ocular diameter (6.4–11.3% of HL versus 4.0– 6.3%), the number of scales above the lateral line at the midbody (11–12 versus eight to nine), and the number of caudal-fin rays (10–16 versus 18–20). Apteronotus galvisi is differentiated from A. rostratus by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), and the number of scales above the lateral line at the midbody (11–12 versus nine to ten). Apteronotus galvisi is distinguished from A. spurrellii by the ocular diameter (6.4–11.3% of HL versus 3.7-5.8%), the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the number of scales above the lateral line at the midbody (11–12 versus eight to nine), the number of anal-fin rays (145–165 versus 171–173), and the condition of the lateral ethmoid (ossified versus cartilaginous).
Description: Head, body, and fin shape and pigmentation illustrated in Figure 11 View Figure 11 . Morphometrics for holotype and paratypes presented in Table 2. Body laterally compressed, greatest body depth located at, or slightly posterior to, abdominal cavity. Dorsal
N, number of specimens; H, holotype. Range includes holotype. Tail depth data unavailable for A. galvisi
profile of body nearly straight. First perforated scale of lateral line located above pectoral-fin origin. Lateral line extending posteriorly to base of caudal fin. Scales above lateral line to mid-dorsal line at midbody 11(11) or 12*(4).
Head laterally compressed, widest at opercular region and deepest at nape. Dorsal profile of snout and overall head nearly straight with snout longer in mature males than mature females. Eye small, located laterally on head, and completely covered by thin membrane. Anterior naris located at end of small tube and close to tip of snout. Posterior naris ellipsoidal, without tube and positioned closer to tip of snout than to anterior margin of eye. Mouth terminal with rictus extending posterior of vertical through anterior border of eye. Branchial opening located anterior to vertical through pectoral-fin insertion. Anus and urogenital papillae located posterior to vertical through eye without ontogenetic variation in position.
Pectoral fin elongate, with 13*(3), 14(3), 15(7), 16(2), or 17(1) rays. Anal-fin origin located anterior to vertical through posterior margin of opercle. Total anal-fin rays 145(1), 146(2), 147(1), 148(2), 152*(3), 153(1), 155(1), 156(1), 161(1), or 165(2). Tail compressed and short; ending in small, elongate caudal fin. Caudal-fin rays 10 (2), 11(1), 12*(6), 14(2), or 16(1).
Origin of dorsal sagittal electroreceptive filament situated on posterior half of body. Filament inserted into narrow mid-dorsal groove and almost extending to, or slightly beyond, vertical through posterior terminus of anal-fin base.
Secondary sexual dimorphism: Sexually mature males (N = 3, 160– 174 mm TL) and females (N = 9, 124– 189 mm TL) of A. galvisi differ in overall head morphology with the dorsal profile of head straight in mature males versus slightly curved ventrally in immature and mature females ( Fig. 11 View Figure 11 ; de Santana et al., 2007: fig. 1).
Coloration in alcohol: Body and head grey to light brown. Prominent, broad, cream-coloured or yellow band extending from chin along dorsal midline of head and body to beginning of dorsal filament. Pectoral and anal-fin rays brown with inter-radial membranes hyaline. Pale band encircling base of caudal fin. Caudal fin light coloured basally and brown distally.
Distribution: Apteronotus galvisi is known from the Ríos Cusiana, Cravo, Pauto, and Upia in the piedmont of the Cordillera Oriental, 305–424 m asl, in the upper Río Meta system, Colombia ( Fig. 5 View Figure 5 ).
Electric organ discharges (EODs): As is the case in all other recorded species of the Apteronotidae , A. galvisi generates wave-type EOD with frequencies in the species ranging from 700–957 Hz, without overlapping frequencies between mature males and females ( de Santana et al., 2007).
Remarks: The Río Orinoco basin is home to four of the nine species in the A. leptorhynchus species-group. This high percentage correlates with this river system being by far the largest basin within the range of the species-group. Two of the species, A. baniwa and A. pemon , are limited to southern portions of the Orinoco basin with known areas of occurrence distant from both each other and from A. galvisi and A. macrostomus , the two other members of the species-group in the system. Apteronotus galvisi and A. macrostomus , in contrast, both occur within a relatively limited region in the western portion of the Orinoco basin.
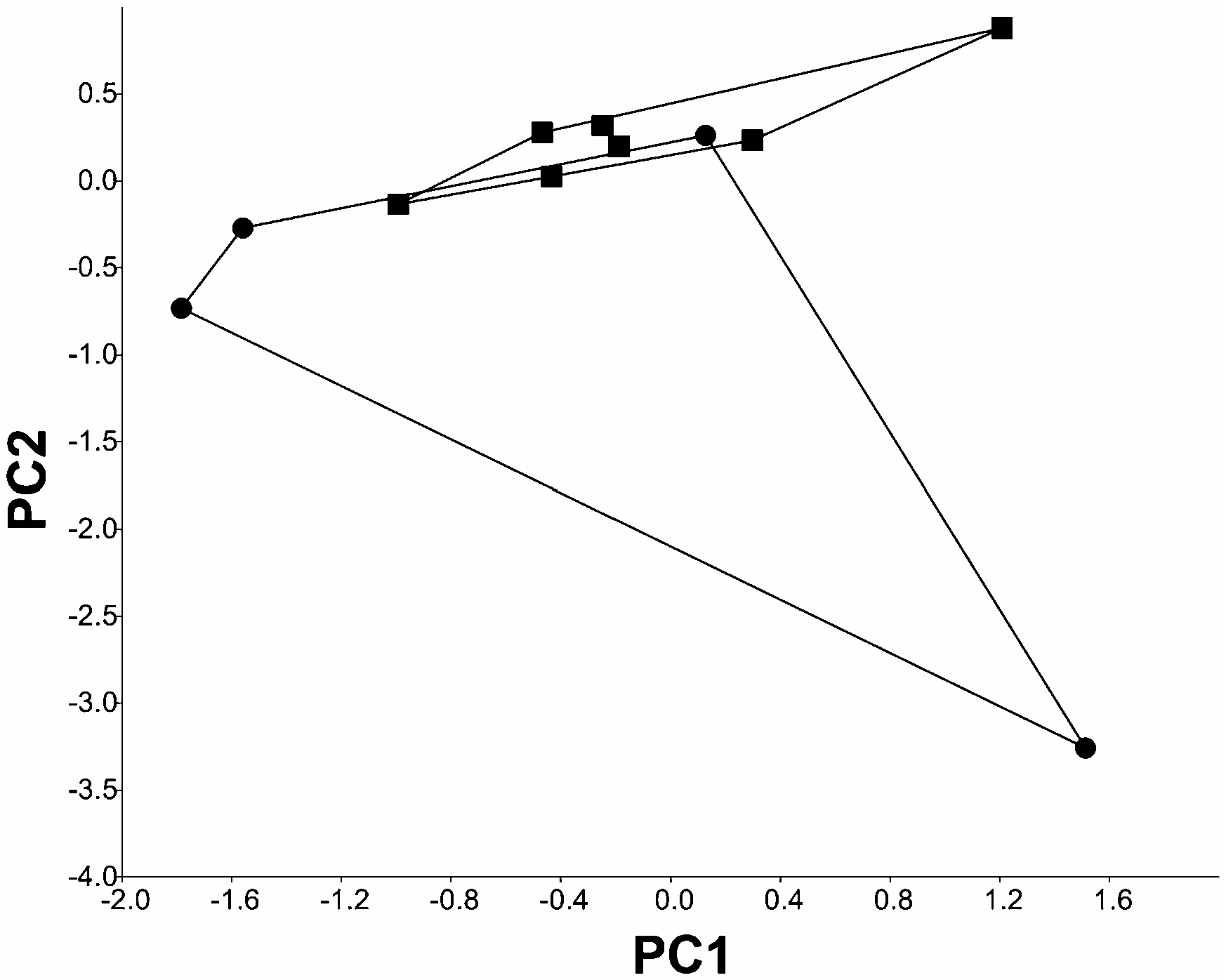
Notwithstanding their geographical proximity, A. galvisi and A. macrostomus have a distinct separation in the ranges of caudal-fin rays (10-16 versus 18-21, respectively); a difference all the more noteworthy because the samples of these two species are the largest available in the A. leptorhynchus speciesgroup. The results of the PCA showed the separation between A. galvisi and A. macrostomus along the second and third principal components ( Fig. 12 View Figure 12 ). PC2 and PC3 reflected 4.6 and 1.4% of the variation, respectively. The best predicators of the differences between the two species were: the internarial dis- tance (0.54), the ocular diameter (0.76), the interorbital distance (0.58), the branchial opening (0.44), and the pectoral fin length (0.35). The jack-knife crossvalidation procedure classified 100% of the specimens between these species, further supporting the recognition of these as valid species. Finally, although mature males of both species are relatively rare, it is noteworthy that mature males of A. galvisi (three specimens, 160–174 mm TL) can be distinguished from the only known mature male of A. macrostomus (the holotype, 265 mm TL) by the head length (18.6– 20.3% of LEA versus 21.4%), the head width (25.3– 36.5% of HL versus 39.3%), the head depth (37.9– 39.4% of HL versus 50.1%), the snout length (40.0– 43.8% of HL versus 44.7%), the distance from the posterior naris to the eye (17.8–20.3% of HL versus 23.6%), the width of the mouth (40.6–50.1% of HL versus 56.2%), the intraocular width (14.1–18.6% of HL versus 19.9%), and the postocular distance (49.2– 58% of HL versus 62.1%). Many of these differences are more pronounced than those between mature males of different species in other groups within the Apteronotidae ; however, additional material is necessary to test whether these differences, or some subset of them, serve to unambiguously delimit the two species.
Material examined: Colombia. Casanare. IAvHP 8133 , holotype, male, 184.4 mm TL; Río Meta drainage, Río Cusiana, at El Venado bridge, 305 m asl, 05°00′50′′N, 72°41′30″W. Paratypes: IAvHP 8130 , two immatures, 97.5–99.2 mm TL; same data as holotype. IAvHP 8129 , three males, 125–174.2 mm TL, three females 132.8–163.3 mm TL; Yopal, Quebrada La Aguatoca at bridge on road to Morro, 357 m asl, 05°26′21″N, 72°27′09″W. IAvHP 8131 , two females, 124.8– 162.6 mm TL; Río Pauto at bridge on road to Yopal- Pore, 306 m asl, 05°33′44′′N, 72°08′44″W. IAvHP 8132 , one immature, 98.3 mm TL, one female, 158.4 mm TL, Río Upia at bridge on road to Sisga, 327 m asl, 04°49′09″N, 73°04′57″W. IAvHP 8167 , three females, 170.6–189.8 mm TL; Yopal, Río Cravo Sur at Colgante bridge, 424 m asl, 05°25′37″N, 72°27′11″W GoogleMaps .
APTERONOTUS LEPTORHYNCHUS (ELLIS)
( FIGS 5 View Figure 5 , 13 View Figure 13 ; TABLE 2)
Sternarchus leptorhynchus Ellis View in CoL in Eigenmann, 1912: 439 [original description; type-locality Amatuk, Guyana]. – Ellis, 1913: 147, pl. 23, fig. 4 [redescription based on Ellis, in Eigenmann, 1912; first illustration of species].
Apteronotus leptorhynchus, Fowler, 1951: 426 View in CoL [comb. nov.; in part, citations from Guyana; not citations from upper Amazon]. – Lasso, Machado-Allison & Pérez Hernández, 1989: 122 [ Essequibo River]. – Mago-Leccia, 1994: 29 [in listing of species; not illustrated specimen from Río Orinoco basin]. – Albert, 2003: 498 [in part; in listing of species; Guyana; not citations from other regions of South America]. – Albert, 2009: 46 [in part; Guyana; not citations from other regions of Guiana Shield]. – de Santana & Cox-Fernandes, 2012: 284 [ Essequibo River basin; occurrence in streams].
Diagnosis: Apteronotus leptorhynchus is differentiated from A. anu by the condition of the fifth infraor- bital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of scales above the lateral line at the midbody (12–13 versus seven to nine), the number of pectoral-fin rays (17–18 versus 14–16), the number of anal-fin rays (151–156 versus 159–176), the number of caudal-fin rays (17–21 versus 10–13), and the number of scales along the lateral line (78–82 versus 59–63). Apteronotus leptorhynchus differs from A. baniwa in the number of scales above the lateral line at the midbody (12–13 versus seven to eight), the number of scales along the lateral line (78–82 versus 61–70), the number of pectoral-fin rays (17–18 versus 14–16), the number of precaudal vertebrae (18 versus 16–17), and the number of total vertebrae (78–81 versus 68–77). Apteronotus leptorhynchus is distinguished from A. ferrarisi by the mouth length (35.1–48.8% of HL versus 49.4–56.3%) and the tail length (17.1–18.6% of LEA versus 6.9-9.9%). Apteronotus leptorhynchus is differentiated from A. galvisi by the number of caudal-fin rays (17–21 versus 10–16), the length of the anal-fin base (81.7–83.7% of LEA versus 69.4– 81.1%), and the tail length (17.1–18.6% of LEA versus 11.8–15.9%). Apteronotus leptorhynchus differs from A. macrostomus in the number of pectoral-fin rays [17–18 (17 pectoral-fin rays present in one of four specimens examined) versus 14–17 (17 pectoral-fin rays present in two of 13 specimens examined)]; in addition, the holotypes of A. leptorhynchus and A. macrostomus , mature males of nearly identical sizes, differ from each other in the head depth (43.6% of HL versus 32.5%), the snout length (41.9% of HL versus 44.7%), the mouth length (48.8% of HL versus 56.2%), and the postorbital distance (57.9% of HL versus 48.7%). Apteronotus leptorhynchus is distinguished from A. pemon by the number of scales above the lateral line at the midbody (12–13 versus eight to nine) and the number of pectoral-fin rays (17–18 versus 16). Apteronotus leptorhynchus is differentiated from A. rostratus by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the number of scales above the lateral line at the midbody (12–13 versus nine to ten), the number of caudal-fin rays (17–21 versus 14–16), the number of scales along the lateral line (78–82 versus 63–67), the tail length (17.1–18.6% of LEA versus 10.6–12.4%), and the condition of the lateral ethmoid (ossified versus cartilaginous). Apteronotus leptorhynchus differs from A. spurrellii in the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of pectoral-fin rays (17–18 versus 12 -13), the number of anal-fin rays (151–156 versus 171–173), the number of scales along the lateral line (78–82 versus 73–76), and the extent of the mid-dorsal groove (extending eight scales beyond the vertical through the posterior terminus of the anal fin versus four scales).
Description: Head, body, and fin shape and pigmentation illustrated in Figure 13 View Figure 13 . Morphometrics for holotype and paratypes presented in Table 2. Body laterally compressed, greatest body depth located at, or slightly posterior to, abdominal cavity. Dorsal profile of body nearly straight. First perforated scale of lateral line located above pectoral-fin origin. Lateral line extending posteriorly to base of caudal fin. Scales along lateral line 78(1), 80(1), or 82*(1). Scales above lateral line to mid-dorsal line at midbody 12*(3) or 13(1).
Head laterally compressed, widest at opercular region and deepest at nape. Dorsal profile of snout and overall head nearly straight. Eye small, located laterally on head, and completely covered by thin membrane. Anterior naris located at end of small tube and close to tip of snout. Posterior naris ellipsoidal, without tube and positioned closer to tip of snout than to anterior margin of eye. Mouth terminal with rictus situated posterior of vertical through anterior margin of eye. Branchial opening located slightly anterior to vertical through pectoral-fin insertion. Anus and urogenital papillae located posterior to vertical through eye and without ontogenetic variation in position.
Pectoral fin elongate, with 17(2) or 18*(2) rays. Anal-fin origin located at, or slightly anterior to, vertical through posterior margin of opercle. Unbranched anal-fin rays 19*(1) or 22(2); total anal-fin rays 151(1), 152*(1), or 156(1). Tail compressed and short; ending in small, elongate caudal fin. Caudal-fin rays 17(1), 19(1), or 21*(1).
Origin of dorsal sagittal electroreceptive filament located on posterior half of body. Filament inserted into narrow mid-dorsal groove extending 8*(2) scales beyond vertical through posterior terminus of anal fin. Precaudal vertebrae 18*(4). Total vertebrae 78(2) or 81*(1).
Coloration in alcohol: Body and head light to dark brown. Prominent, pale band extending from chin along dorsal midline of head and body to beginning of dorsal filament. Pectoral- and anal-fin rays dark brown with inter-radial membranes translucent. Pale band encircling base of caudal fin. Caudal fin pale at base and dark brown posteriorly.
Distribution: Apteronotus leptorhynchus is known only from the Essequibo River drainage in Guyana ( Fig. 5 View Figure 5 ).
Remarks: Apteronotus leptorhynchus has been reported from broad regions of northern South America (cis- and trans-Andean regions of Colombia – Mojica-C, 1999: 563; Lago Maracaibo – Mago-Leccia, 1970: 77; Taphorn et al., 1997: 80; Río Caura, Rodríguez-Olarte et al., 2003: 198; Lasso et al., 2003: 238; Lasso et al., 2010: 70; Río Orinoco basin and Río Paria – Cala, 1977: 10; Lasso et al., 2004a: 181, 2004b: 142; Pacific and Caribbean drainages of Colombia – Maldonado-Ocampo et al., 2008: 213; Peru – Fowler, 1945: 182, 1951: 426; and Venezuela – Lasso et al., 2004a: 181). Available information, however, indicates that A. leptorhynchus is endemic to the parts of the Essequibo River basin in Guyana. The above records are either misidentifications of specimens of other species-groups or may be based on species described as new herein. Lasso et al. (2010: 70) reported A. leptorhynchus from the Río San Juan, a river draining into the Golfo de Paria north of the mouth of the Río Orinoco. Owing to lack of access to the voucher specimen we were unable to evaluate this record.
Although the primary diagnostic feature discriminating A. leptorhynchus from A. macrostomus , the number of pectoral-fin rays, shows a limited degree of overlap in the available samples of the species, the holotypes of the two species, both mature males of approximately the same body size, show trenchant differences in various details (see Diagnosis for each species). In addition, various features of mature males not readily expressed via standard meristics and morphometrics differ notably between mature males of the two species. Most obvious of these is the form of the snout and dorsal surface of the head, which is straight in the case of A. leptorhynchus versus notably convex in A. macrostomus (see Figs 13 View Figure 13 , 15 View Figure 15 ). Furthermore, the results of the PCA showed partial notable, albeit not absolute, separation between A. leptorhynchus and A. macrostomus along the first and second principal components ( Fig. 14 View Figure 14 ). PC1 and PC2 reflected 91.6 and 5.8% of the variation, respectively. The best predicators of the differences between the two species were: the distance from the posterior snout to the eye (0.36) and the head depth (-0.94). A jack-knife cross-validation procedure correctly classified 86.6% of the specimens by species, further supporting the hypothesis of the differences between the two species. Those morphological differences, supplemented by the distinct gap in the known distributions of the two species, support the recognition of the two nominal forms as valid.
Material examined: Guyana. BMNH 1974.5 .22.136, 1, 124; Potaro River . CAS 62333, 1 About CAS , 100 About CAS ; Essequibo River , Potaro River at Amatuk, 1908. FMNH 53294 About FMNH , 1 About FMNH , 260 About FMNH ; holotype, Amatuk, 1908. FMNH 53295 About FMNH , 1 About FMNH , 95 About FMNH , Warraputa, 1908. INHS 49524 About INHS , 1 About INHS , 94 About INHS ; Potaro River , Essequibo River basin, Amatuk Cataract, Macaroni- Potaro 05°18′13″N, 59°18′40″W GoogleMaps .
APTERONOTUS MACROSTOMUS (FOWLER)
( FIGS 15 View Figure 15 , 16 View Figure 16 ; TABLE 2)
Sternarchus macrostomus Fowler, 1943: 263 View in CoL , fig. 63 [original description; type-locality: Villavicencio, Río Meta basin, Colombia].
Apteronotus macrostomus, Mago-Leccia, 1994: 30 View in CoL , fig. 35 [comb. nov.; in listing of species]. – Albert, 2003: 499 [in listing of species; Río Meta basin, Colombia]. – Lasso et al., 2004a: 181 [Orinoco basin]. – Lasso et al., 2004b: 142 [western Orinoco basin]. – de Santana & Cox-Fernandes, 2012: 284 [Río Orinoco basin; occurrence in rivers].
‘Apteronotus’ macrostomus, Albert View in CoL & Campos-da- Paz, 1998: 431 [phylogenetic position].
Diagnosis: Apteronotus macrostomus is distinguished from A. anu by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of anal-fin rays (140–152 versus 159–176), the number of caudalfin rays (18–21 versus 10–13), and the number of scales above the lateral line at the midbody (11–14 versus seven to nine). Apteronotus macrostomus is differentiated from A. baniwa by the number of scales above the lateral line at the midbody (11–14 versus seven to eight), the number of caudal-fin rays (18–21 versus 14–17), and the number of precaudal vertebrae (18 versus 16–17). Apteronotus macrostomus differs from A. ferrarisi in the tail length (13.9–20.6% of LEA versus 6.9-9.9%), the number of caudal-fin rays (18–21 versus 15–17), and the number of total vertebrae (75 versus 61–71). Apteronotus macrostomus is distin- guished from A. galvisi by the number of caudal-fin rays (18–21 versus 10–16). Apteronotus macrostomus is differentiated from A. leptorhynchus by the number of pectoral-fin rays [14–17 (17 pectoral-fin rays present in two of 13 specimens examined) versus 17–18 (17 pectoral-fin rays present in one of four specimens examined)]; in addition, the holotypes of A. macrostomus and A. leptorhynchus , both mature males of nearly identical sizes, differ from each other in the head depth (32.5% of HL versus 43.6%), the snout length (44.7% of HL versus 41.9%), the mouth length (56.2% of HL versus 48.8%), and the postorbital distance (48.7% of HL versus 57.9%). Apteronotus macrostomus is distinguished from A. pemon by the number of scales above the lateral line at the midbody (11–14 versus eight to nine). Apteronotus macrostomus is distinguished from A. rostratus by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of scales above the lateral line at the midbody (11–14 versus nine to ten), the number of anal-fin rays (140–152 versus 153–162), the number of caudal-fin rays (18–21 versus 14–16), and the tail length (13.9– 20.6% of LEA versus 10.6–12.4%). Apteronotus macrostomus is differentiated from A. spurrellii by the condition of the fifth infraorbital (present as a tubular laterosensory canal segment versus absent), the condition of the lateral ethmoid (ossified versus cartilaginous), the number of pectoral-fin rays (15–17 versus 12–13), the number of scales above the lateral line at the midbody (11–14 versus eight to nine), the number of anal-fin rays (140–152 versus 171–173), and the number of caudal-fin rays (18–21 versus 14).
Description: Head, body, and fin shape and pigmentation illustrated in Figure 15 View Figure 15 . Morphometrics for holotype and paratypes presented in Table 2. Body laterally compressed with greatest body depth located at, or slightly posterior to, abdominal cavity. Dorsal profile of body nearly straight. First perforated scale of lateral line located above pectoral-fin origin. Lateral line extending posteriorly to base of caudal fin. Scales along lateral line 79(3), 80(4), or 81*(5). Scales above lateral line to mid-dorsal line at midbody 11(5), 12*(3), 13(1), or 14(3).
Head laterally compressed, widest at opercular region and deepest at nape. Dorsal profile of head nearly straight. Eye small, located laterally on head, and completely covered by thin membrane. Anterior naris located at end of small tube and close to tip of snout. Posterior naris ellipsoidal, without tube and positioned closer to tip of snout than to anterior margin of eye. Mouth terminal with rictus extending posterior of vertical through anterior border of eye. Branchial opening located slightly anterior to vertical through pectoral-fin insertion. Anus and urogenital papillae located posterior to vertical through eye without ontogenetic variation in position.
Pectoral fin elongate, with 14*(3), 15(2), 16(2), or 17(5) rays. Anal-fin origin located at, or slightly anterior to, vertical through posterior margin of opercle. Unbranched anal-fin rays 19*(1), 22(4), 23(2), 25(3), or 27(2); total anal-fin rays 140(1), 141(1), 143(2), 146*(3), 148(1), or 152(1). Tail compressed and short; ending in small, elongate caudal fin. Caudal-fin rays 18(3), 19(4), or 21*(3).
Origin of dorsal sagittal electroreceptive filament located on posterior half of body. Filament inserted into narrow mid-dorsal groove ranging from slightly anterior to slightly posterior of vertical through posterior terminus of anal fin. Precaudal vertebrae17(3) or 18*(2). Total vertebrae 71(1), 73(2), 74(2), or 75(1).
Coloration in alcohol: Body and head lightly coloured to dark brown. Pale, broad, longitudinal band extending from chin along dorsal midline of head and body to beginning of dorsal filament. Pectoral- and anal-fin rays dark brown with inter-radial membranes translucent. Pale band encircling base of caudal fin. Caudal fin pale basally and dark brown distally.
Distribution: Apteronotus macrostomus is known from the Río Meta at Villavicencio, Colombia ( Fig. 16 View Figure 16 ).
Remarks: See under Remarks for A. galvisi concerning differences between that species and A. macrostomus and the diagnostic features discriminating the single available mature male (holotype) of A. macrostomus from mature males of A. galvisi . Features distinguishing A. galvisi from A. leptorhynchus are discussed in the Remarks for the latter species.
Material examined: Colombia. Meta. ANSP 70528, 1, holotype, 265; Río Meta at Villavicencio (4°08′S, 73°40′W). CAS [ SU 23728 About CAS ], 1, 205; Guaicaramo. FMNH 92638 About FMNH , 8 About FMNH , 83–145 About FMNH ; Caño Negro on road to Puerto Porfia , east of Villavicencio. FMNH 92715 About FMNH , 1 About FMNH , 85 About FMNH , Campo Alegre , Campo Aguasclaras east of Villavicencio. USNM 100808 About USNM , 1 About USNM , 172 About USNM , Villavicencio , Río GoogleMaps Meta basin. UF 33218, 1, 157; Río Meta, Caño Quenane (4°02′S, 73°10′W). UF 36604, 5, 71–230, Río Guatiguia, near Villavicencio GoogleMaps .







| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Apteronotus ferrarisi
| Santana, Carlos David De & Vari, Richard P. 2013 |
Apteronotus galvisi
| de Santana CD & Cox-Fernandes C 2012: 284 |
| Machado-Allison A & Lasso CA & Usma JS & Sanchez-Duarte P & Lasso-Alcala OM 2010: 242 |
| Urbano-Bonilla A & Zamudio J & Maldonado-Ocampo JA & Bogota-Gregory JD & Cortes-Millan GC & Lopez Y 2009: 162 |
| Maldonado-Ocampo JA & Vari RP & Usma JS 2008: 213 |
| de Santana CD & Maldonado-Ocampo J & Crampton WGR 2007: 118 |
Apteronotus macrostomus, Mago-Leccia, 1994: 30
| de Santana CD & Cox-Fernandes C 2012: 284 |
| Lasso CA & Lew D & Taphorn DC & DoNascimiento C & Alcala O & Provenzano F & Machado-Allison A 2004: 181 |
| Lasso CA & Mojica JI & Usma JS & Maldonado JA & DoNascimiento C & Taphorn DC & Provenzano F & Lasso-Alcala OM & Galvis G & Vasquez L & Lugo M & Machado-Allison A & Royero R & Suarez C & Ortega-Lara A 2004: 142 |
| Albert JS 2003: 499 |
| Mago-Leccia F 1994: 30 |
Apteronotus leptorhynchus, Fowler, 1951: 426
| de Santana CD & Cox-Fernandes C 2012: 284 |
| Albert JS 2009: 46 |
| Albert JS 2003: 498 |
| Mago-Leccia F 1994: 29 |
| Lasso CA & Machado-Allison A & Perez Hernandez R 1989: 122 |
| Fowler HW 1951: 426 |
Sternarchus macrostomus
| Fowler HW 1943: 263 |