
Zophopetes dysmephila Trimen, 1868
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3831.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:2EF9A3DB-0EAA-4384-8ADA-A7D269E5904D |
|
DOI |
https://doi.org/10.5281/zenodo.5121694 |
|
persistent identifier |
https://treatment.plazi.org/id/6F3587EC-3228-1B5B-AB9F-59EDFE74E183 |
|
treatment provided by |
Felipe |
|
scientific name |
Zophopetes dysmephila Trimen, 1868 |
| status |
|
Zophopetes dysmephila Trimen, 1868 View in CoL
This sexually dimorphic species was described as Pamphila dysmephila by Trimen (1868) from South Africa (type locality ‘Bashee River, Kaffraria’, i.e. Mbhashe River, Eastern Cape Province) based on a female. The male was subsequently described as Hesperia schulzi Plötz (1882) from Angola and as Hesperia mucorea Karsch (1892) from Cameroon. Evans (1937) treated these two names as synonyms of Z. dysmephila , and this view has not been questioned. In preparing this paper, it became apparent that there is geographical variation in the caterpillars and male genitalia, suggesting either a complex species or a species complex.
The first illustration of the male genitalia is that of Murray (1959) based on a specimen from Port Elizabeth (Eastern Cape Province, and hence representative of the type locality), which shows more or less symmetrical tapered, pointed valves. Examination of the BMNH collection suggests that W.H. Evans examined the male genitalia of just one specimen, which was from ‘Zululand’ (largely overlapping with KwaZulu-Natal Province, north of the type locality). Although Evans (1937) shows only one valve in his diagram, his preparation shows that the valves are asymmetrical, the left valve resembling that shown in Evans’ figure, but the right more like those shown by Murray (1959). Larsen (2005, Figure 7.21 View FIGURE 7 ) illustrates the right valve of a Nairobi specimen, which has a strongly upturned apical portion, differing from the earlier illustrations.
In preparing this paper, MJWC compared the dissected male genitalia of single Kenyan specimens from Kakamega Forest (western Kenya, west of the Rift Valley ), Nairobi ( central Highlands , east of the Rift Valley) and Bamburi at the coast. The Nairobi specimen was clearly very different from the other two, resembling Larsen’s (2005) figure, with the apical portion of the valves turned upwards and inwards. Brushing the scales off the ventral surface of male genitalia, particularly if the genitalia are extruded when preparing fresh specimens, will reveal this character in many pinned specimens and showed that all six other MJWC specimens from Nairobi have valves of this type, as does one from Nyeri, Central Highlands, Kenya, in the Hope Entomological Collections, Oxford University Museum ( HEC). This form is referred to as ‘ Nairobi typical’ below. The specimens from Kakamega Forest and Bamburi are similar to each other, but still show potentially significant differences, e.g. the right valve of the Bamburi specimen is more elongate with a distinctly rounded apex, compared to the pointed apex of the specimen from Kakamega Forest. A male from Eldoret, western Kenya, in HEC probably matches those from Kakamega Forest —certainly it is not of the Nairobi form. A male from Eritrea in the BMNH has the male genitalia displayed in situ and appears to match the Bamburi male. However, T. B. Larsen (pers. comm. 2013) has dissected males from various localities and finds substantial variation and no clear pattern. Thus, apart from the series from Nairobi, no material in series has been examined from any localities, and although the male genitalia of the Nairobi form seem constant, this may not be the case elsewhere. As yet no female genitalia have been documented. The taxonomy of this ‘species’ needs further research. Here, we provide information on the early stages as we and others have observed them in different parts of the range of Z. dysmephila .
The main population stretches from the coastal areas of South Africa, as far west as Western Cape Province ( Henning et al. 1997) to most of eastern Africa, north to Kenya and Uganda, but not much west of the Albertine Rift, although MJWC found early stages at Butare, Rwanda (90/202). It seems commonest in South Africa, but as indicated below, its apparent general scarcity may be more apparent than real. The type localities of the synonyms schulzi from Angola and mucorea from Cameroon lie outside this area, which as T.B. Larsen (pers. comm. 2013) points out, is puzzling. There are three or four specimens of typical dysmephila from Bom Jesus, Angola which might well have been imported with palms from Mozambique (T.B. Larsen pers. comm. 2013). Indeed, the movement of ornamental Phoenix spp. is not rare and populations of Z. dysmephila may have been redistributed with them, both within and occasionally between countries.
There is a population in Eritrea ( Evans 1947, Jannone 1948), which might have been introduced (C.B. Williams in Jannone 1948), but a more recent record from Bonga, near Jimma, Ethiopia (ABRI collection) suggests the Eritrean population may not be so isolated. The record from Eritrea is probably the basis of Gentry’s (1965) record of Z. dysmephila from Ethiopia, since Eritrea was not separated at that time.
In Kenya and Tanzania this species is probably to be found wherever its main food plants, Phoenix spp. palms, grow (or are grown as ornamentals). It is particularly common in Nairobi where many P. reclinata have been planted in gardens and public places, and many of these will have at least one leaf shelter, often several.
Adult behaviour. Adults are seldom seen. They are reported to fly at dusk ( Trimen 1889, Pinhey 1965), making a loud humming noise with their wings (J.H. Bowker in Trimen 1889) and come to flowers, particularly those of papaya. Dickson & Kroon (1978) also report that they will fly on warm wet days or very late in the afternoon. At dusk they fly rapidly around the crown of palm trees according to Kielland (1990), and TCEC caught specimens flying round a bougainvillea bush at dusk in a Dar es Salaam garden. MJWC recently observed and photographed a male flying around a clump of P. reclinata View in CoL at 16.30h (Gigiri, Nairobi, Oct 2013), and has caught a male in a mercury vapour light trap used for catching moths in his Nairobi garden ( Figure 3.1 View FIGURE 3 ). In contrast, the early stages are easy to locate where the food plants are found.
The margin of the apical part of the male forewing upper surface is white, an unusual feature in Hesperiidae , but one shared with Carystoides a Neotropical genus where again only males show it ( Evans 1955, Cock 2005), and some species of the Oriental genus Erionota , e.g. E. hiraca (Moore) ( Swinhoe 1912 –1913, Evans 1949 as E. acroleucus (Wood-Mason & De Nicéville)) and E. surprisa De Jong & Treadaway (1993) . The white band of the antenna club is also conspicuous and reminiscent of the white antennae of Chondrolepis spp. , which are pure white above in the male and have the club partly or completely white above in the female ( Evans 1937, De Jong 1986). All four genera fly at dusk and this colouring may well be a signalling device, for mate location and acceptance, as suggested by De Jong (1986) for Chondrolepis .
Food plants. This is one of the first skipper life histories to have been documented from Africa. Trimen (1889) reports J.H. Bowker’s 1881 observations of the caterpillar, pupa and food plant from South Africa, based on A.D. Millar’s discoveries at Durban (=D’Urban). The food plant reported is the indigenous Phoenix reclinata grown as a garden plant. It is not clear whether subsequent records of P. reclinata from other countries represent local observations, independently reporting this food plant, or simply repeat the earlier information from South Africa. However, locally attributable records from Uganda ( Le Pelley 1959), Kenya (MJWC, SCC), Tanzania (TCEC) and Zambia (TCEC) suggest that P. reclinata is a normal food plant throughout this skipper’s range, and this probably represents the original food plant.
Other species of Phoenix are also reported as food plants. The introduced ornamental P. canariensis is a suitable food plant, and Jannone (1948) reported Z. dysmephila as a pest on this palm in Eritrea, where he considered it to be the preferred food plant (la specie … che viene particolarmente presa di mira). An introduced population has also been reported on this palm in Cape Town ( Geertsema 1985) although a subsequent report by Claassens & Dickson (1986) referred to date palms or common date palms––by implication P. dactylifera , although this was not made explicit. MJWC found caterpillars on a palm provisionally identified as P. canariensis at Diani Beach, Kenya (95/102).
Dickson & Kroon (1978) report that C.G. Clark ‘frequently obtained larvae from the leaves of the common date palm, P. dactylifera ’ at Port Elizabeth, South Africa, whereas Pringle et al. (1994) refer to the common date palm, without a scientific name, and then list the food plants as P. ‘ reclinata (wild date palm)’ and P. ‘ dactylifera (date palm)’. Jannone (1948) reported that in Eritrea, in the proximity of infested P. canariensis , the palms Chamaerops humilis (a Mediterranean species) and P. dactylifera are also frequently attacked (riscontrandosi frequentemente nelle prossimità di Phoenix canariensis più o meno infestate dalla specie in questione, esemplari inquinati e danneggiati di Chamaerops humilis e Phoenix dactylifera ). In a similar observation, A.J.M. Claassens found caterpillars in Cape Town ‘on P. canariensis and another palm, tentatively identified as Chrysalidocarpus (Areca) lutescens [i.e. Dypsis lutescens ], the butterfly palm, standing between specimens of P. canariensis ’ ( Claassens & Dickson 1986) . MJWC recently found empty shelters ( Figure 4.1 View FIGURE 4 ) and feeding on P. reclinata ( Figure 4.2 View FIGURE 4 ), P. sylvestris (an Indian species, Figure 4.3 View FIGURE 4 ) and Chamaerops humilis ( Figure 4.4 View FIGURE 4 ) planted on the campus of the World Agroforestry Centre (ICRAF), Gigiri, Nairobi (15 Nov 2013, MJWC 13/03–04), which can be attributed to Z. dysmephila , as no other palm-feeding skippers occur in the Nairobi area.
In addition to Phoenix spp. , Van Someren (1974) lists Raphia spp. and Borassus spp. (Arecaceae) as food plants for East Africa. Probably the latter two can be referred to R. farinifera (raffia palm) and B. aethiopum (borassus or African fan palm), the only indigenous members of these genera in Kenya ( Beentje 1994). Sevastopulo (1975) lists Raphia , but did not rear this species himself (Sevastopulo unpublished). These food plants have been repeated by subsequent authors. However, in Tanzania TCEC has confirmed R. farinifera to be a food plant, but not B. aethiopum . Although TCEC has collected caterpillars from B. aethiopum , he has never found any Hesperiidae on it. Accordingly we consider the record from Borassus likely to be an error.
As discussed under Zophopetes above and Z. cerymica below, early records from French West Africa of Z. dysmephila as a pest of oil palm and coconut ( Genty 1968, Mariau & Morin 1974) are misidentifications for Z. cerymica , which have been repeated in the economic literature (e.g. Lever 1969, Howard & Abad 2001). Larsen (1991) includes Cocos (i.e. C. nucifera , the only species of the genus) as a food plant of Z. dysmephila . We have not found Z. dysmephila feeding on coconut in Kenya or elsewhere.
To summarise, the original food plant is most probably the indigenous P. reclinata . Raphia farinifera is used as a food plant, but we do not know if this behaviour is widespread or common. The introduced P. canariensis is an acceptable food plant, but it is not clear how regularly the introduced P. dactylifera and P. sylvestris are used when available. Carter’s (1992) observation that the caterpillar ‘feeds on date palm ( Phoenix dactylifera ) and other palms’ is therefore slightly misleading. The records on Chamaerops humilis and D. lutescens may represent spillover effects to less-preferred or marginal hosts from populations on Phoenix spp. Records from Cocos nucifera and Borassus sp (p). are not accepted at this time. The suitability of date palm, P. dactylifera , should be further evaluated, since if this important crop is readily accepted, Z. dysmephila would be a significant quarantine pest for the date-growing areas of the world.
Life history. Not only was Z. dysmephila one of the earliest African skippers to have its food plant documented, it was also one of the first to have its life history documented and illustrated. Trimen (1889) included a brief description of the caterpillar, pupa and shelters. A few years later, Kelley (1912) described and illustrated the caterpillar and pupa on P. reclinata at Pietermaritzburg (=Maritzburg), KwaZulu-Natal Province, South Africa. Jannone’s (1948) account of this species as a pest of P. canariensis in Eritrea includes good drawings of all stages, shelters and damage. These two accounts are synthesised briefly by Lepesme (1947). G.C. Clark (in Dickson & Kroon 1978) provided his usual excellent detailed paintings of the complete life history. Here we document the history from Kenya, supported by observations from Rwanda, Tanzania and Zambia.
Ovum. MJWC found the ovum of the Nairobi population is dark red when newly laid and turns white; it is hemispherical 1.7mm wide (range 1.61–1.76, n=7) x 1.1mm high (range 1.06–1.18, n=4), with 27–29 fine ribs which peter out close to the flattened micropyle. One ovum from Kakamega Forest, Kenya, measured 1.57 x 0.98mm wide x high. Ova found at Butare, Rwanda measured 1.6mm wide (range 1.49–1.65, n=21) x 1.0mm high (range 0.98–1.06, n=5), with 34–38 (n=3) fine ribs. In Eritrea, Jannone (1948) reports the ova to be 1.4–1.7 x 1.0mm wide x high.
Ova are laid on palm leaflets, usually singly, on the upper surface near the apex, or in the upper surface hollow at the base of the leaflet. MJWC found that normally ova are laid singly or in pairs, although once a group of four ova was found at Nairobi (90/24) and a group of 24 at Butare, Rwanda (90/202). In Eritrea, Jannone (1948) found that ova are normally laid singly or in groups of up to six, but occasionally groups of up to 40 may be laid; this could be a reflection of high skipper densities. Dead ova are quite commonly found, and many of these have emergence holes of egg parasitoids (below). Ova took 20–30 days to develop at 2300m in Eritrea ( Jannone 1948).
Leaf shelters, Nairobi typical. The newly hatched caterpillar eats the shell of the ovum, apart from the base, and then pulls together the edges of a leaflet towards the apex to form a small channel in which it hides ( Figure 5.2 View FIGURE 5 ). It seems likely that there is considerable predation of first instar caterpillars by spiders and wasps due to the inadequate protection provided by this shelter; certainly hatched ova with no associated shelter, and empty first shelters with no nearby subsequent shelters are quite common. The caterpillar feeds basally or distally to the shelter before making a new shelter lower down the leaf, which it is then able to close to provide much better protection ( Figure 5.4 View FIGURE 5 ). Mature caterpillars will shelter between two leaflets held by silk, and pupate between several leaflets of a young leaf drawn together.
Caterpillar. Clark (in Dickson & Kroon 1978) indicates rearing through six instars, but in Nairobi MJWC documented only five instars (90/24). Below, we segregate the available information by locality.
Caterpillar, South Africa. Kelley (1912), Clark (in Dickson & Kroon 1978, plate 29) and Henning et al. (1997) have documented the early stages of Z. dysmephila View in CoL in southern Africa.
Kelley’s account includes black and white photographs of the caterpillar and pupa from KwaZulu-Natal Province, South Africa. These are not very clear, but the caterpillar can be seen to have a pale head, with the epicranial suture dark and black lines adjacent to the epicranial suture from the vertex, and laterally from vertex to stemmata. There are pale subdorsal lines on the body. We note that the lines adjacent to the epicranial suture extend to the vertex, and seem more or less straight, with little obvious divergence at the adfrontals, although the head is positioned so that this is not clear. Thus, the photo resembles Clark’s (in Dickson & Kroon 1978) figure of the penultimate instar 5, rather than the final instar.
Clark (in Dickson & Kroon 1978) provides a full series of paintings of the life history at Port Elizabeth, Eastern Cape Province, South Africa. The first instar is pale with a dark head. The second instar is pale green and the dark head has a trace of a pale area lateral to the adfrontals. Instar 3 is green with a dark head, with the adfrontals and a streak lateral to them pale. Instar 4 has a dark green dorsal line and the head is now pale, with dark sutures, a dark line adjacent to the epicranial suture and a strong lateral line from the vertex to stemmata, which may extend to the posterior margin. Instar 5 is similar, but the body now has pale subdorsal lines. In the final instar 6 the head and body are paler; the lines adjacent to the epicranial suture start well below the vertex, and diverge sharply along the adfrontal sutures, ending in a slight swelling; the lateral line from vertex to stemmata is broad and appears to extend to the posterior margin; the sutures are dark, and in one of two illustrations, the upper part of the adfrontals is dark.
Henning et al. (1997) provide similar notes on the different instars, as well as a colour photo of the final instar caterpillar, of unattributed origin. In this photograph the subdorsal lines are less distinct, and this is probably a fully mature caterpillar changing to the prepupal colouring, similar to that shown in Figure 20.6 View FIGURE 20 for Z. cerymica View in CoL . The head is similar to that shown by Clark for the final instar, except that the lateral line is almost absent and appears to be represented by a diffuse slightly darker brown area, in this regard matching the head of Nairobi, central Tanzania and Zambia caterpillars ( Figures 7 View FIGURE 7 , 11 View FIGURE 11 , 13 View FIGURE 13 ). Thus it is unclear whether the caterpillar illustrated by Henning et al. (1997) indicates variation in the South African material beyond that documented by Clark (in Dickson & Kroon 1978), or is representative of material from further north in their geographical coverage, e.g. Zimbabwe.
Caterpillar, Eritrea. Jannone (1948) has provided a description and drawings of the caterpillar of Z. dysmephila in Eritrea (which may be indigenous or introduced). His figure 3.2 shows a mature caterpillar in dorsal view with a dark dorsal line but no clear subdorsal lines. The head has black lines adjacent to the epicranial and adfrontal sutures very similar to those illustrated by Henning et al. (1997) and we report below from Zambia. Jannone adds that there is a black band on the posterior margin of the head (una stretta fascia nera in corrispondenza del foro occipitale), and the dorsal part of the adfrontals is black in some specimens ( La zona triangolare compresa tra i due rami anteriori della sutura metopica, in qualche esemplare può essere anche nera). The caterpillar develops in 30–55 days at 2300m in Eritrea depending on the season ( Jannone 1948).
Caterpillar, Nairobi typical. MJWC reared this form several times, and found it to be variable in the head markings, and in the timing of their development in instars 2–4. This is described in some detail to set the observations from other locations in context. Instar 1 (87/6) has a black head, shiny, slightly rugose, 1.0mm wide (range 0.98–1.10, n=6) x 1.1mm wide (range 1.06–1.18, n=6); narrow, black pronotum; body dull shiny green; A8 slightly and A9 quite strongly tinted red; long erect white setae on anal plate. Figure 6 View FIGURE 6 shows a first instar caterpillar about to moult. Instar 2 (87/6) similar; head dark, 1.3mm wide (range 1.25–1.37, n=4) x 1.4mm high (range 1.37–1.49, n=4); body more yellow-green, anal colouring reduced to yellow tint on A9, and white setae on anal plate not obvious. From the second instar, caterpillars may begin to show characteristics of the head markings of the final instar, although the markings are darker and more diffuse. However, in some individuals the head remains mostly black until the fourth (penultimate) instar. Thus, although the head of caterpillars of instar 2 are normally dark, one of two caterpillars collected at Thika (87/6) had the ground colour pale brown; epicranial and clypeal sutures dark; weak diffuse dark line parallel to epicranial suture from well below vertex to and merges into a darker ventral area; diffuse dark band from vertex to stemmata; posterior margin narrowly black.
Instar 3 head 1.75 (range 1.65–1.84, n=13) x 2.05mm high (range 1.88–2.16, n=13). Of the 13 individuals examined, two had a dark head, without distinguishable markings, while at the other extreme, one individual had the head pale brown; epicranial and clypeal sutures black; a diffuse brown line parallel and close to epicranial suture, from well below the vertex to the top of the adfrontals; vertex to apices diffusely brown; posterior margin narrowly dark. There was continuous variation between these extremes, with most caterpillars having the line adjacent to the epicranial suture longer and darker, extending half way down the level of the frons, and a dark band from vertex to apices and stemmata.
Instar 4 head 2.4mm wide (range 2.24–2.71, n=11) x 2.8mm high (range 2.71–2.94, n=11); variable markings. At one extreme, one of 11 caterpillars had the ground colour brown (as opposed to light or pale brown); epicranial, adfrontal and clypeal sutures blackish; dark line adjacent to epicranial suture fused with the dark epicranial suture in lower part, widening and becoming more diffuse to the level of the bottom of the frons; adfrontals pale brown; a wide, dark lateral band from vertex, reaching the posterior margin laterally; posterior margin black. At the other extreme, some caterpillars had markings similar to those of the final instar: ground colour light brown; epicranial and clypeal sutures black; adfrontal suture very slightly darkened in dorsal half; black line adjacent to epicranial suture runs from well below the vertex to the top of the adfrontals; no trace of dark spot on edge of epicranium adjacent to adfrontals; slightly dark, diffuse, lateral band from vertex to apex to half way down side of head; posterior margin narrowly dark. More commonly the dorsal parts of the adfrontals and frons may be dark; there may be a spot on the epicranium, adjacent to the adfrontals, level with half way down the frons; the lateral band may continue to the stemmata.
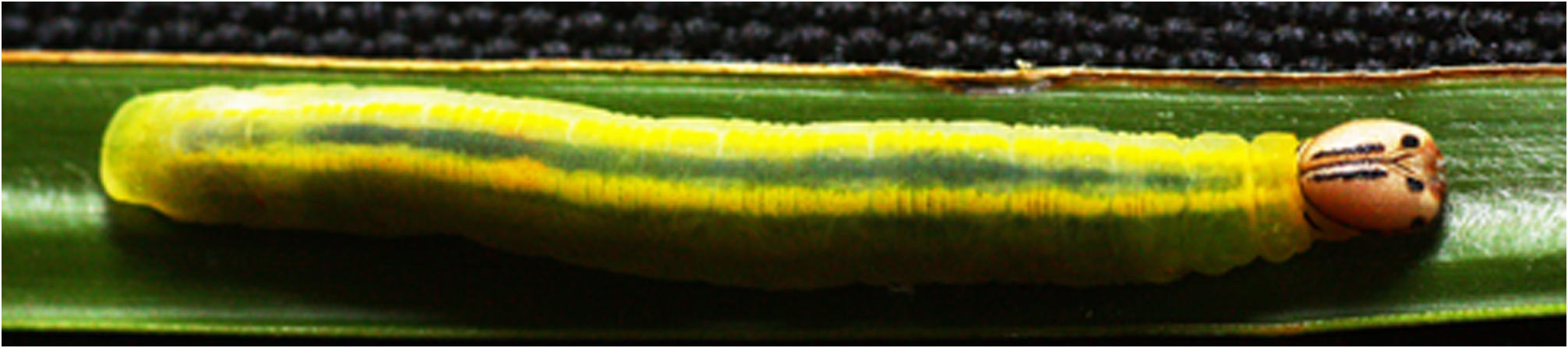
The head of the final, fifth instar caterpillar measures 3.3 x 4.2mm wide x high, and the head markings are variable although less so that in the previous two instars. The following description of the final instar (87/2) is based on one of the first skipper caterpillars that MJWC reared in Nairobi, and represents the typical form––i.e. the form most frequently found around Nairobi, similar to, but not matching those documented from South Africa (above). Length 48mm when collected, six days before it stopped feeding prior to pupation; width 6mm; height 4mm. Other individuals of at least 50mm have been measured (89/35). Head shiny and slightly crinkly; ground colour whitish brown; ventral part of face and adfrontals slightly paler; clypeus pale grey-brown; epicranial suture slightly darker; narrow black line from just below vertex, near and parallel to epicranial suture, diverging slightly and thickening at the level of the top of the adfrontals where it stops; half way from the end of this line to the mouthparts a black spot on the edge of the epicranium, adjacent to adfrontals; stemmata black. The head is held with the mouthparts directed anteriorly (semiprognathous), and the dorsal part resting on top of T1, which has a narrow black pronotum. Body dull green, T2 yellowish green; dark green transparent dorsal line about 1mm wide T3–A7; separated by a narrow line of ground colour, a slightly diffuse dorsolateral line T2–A8; anal plate semicircular, nearly flat, somewhat shiny; spiracle T1 brown, other spiracles pale brown; all legs concolorous. As can be seen in Figure 7 View FIGURE 7 , the intensity of the line adjacent to the epicranial suture is variable, as is the size and intensity of the lower spot adjacent to the adfrontals, but none have been found where the line and spot are joined; similarly, the ground colour varies in intensity, there may be a trace of a lateral band, and the posterior margin may be dark.
Prior to pupation, the bodies become more opaque and yellowish and the dorsal line is more pronounced. The wax glands developed ventrally on A1–A8, covering the whole ventral surface apart from a narrow ventral line, the prolegs, and the posterior half of A7 and A8 (MJWC 89/105). In other caterpillars (MJWC 90/69, 95/102) the wax glands extended to whole ventral area A1–A8 apart from the prolegs.
The young caterpillars feed only from the edge of the leaflet, but mature caterpillars are able to eat through the mid-rib as well, and leave the tip of the leaflet truncated, coming to a point at or near the mid-rib. This is quite characteristic of palm feeding skippers ( Figures 4.1 View FIGURE 4 , 5.5 View FIGURE 5 ), and MJWC has noted it in several other genera around the tropics.
Total caterpillar development takes about 48–57 days in Nairobi. Instars durations of 7, 7, 7, and 7–8 were noted for the first four instars.
Caterpillar, Nairobi atypical. All the caterpillars reared from Nairobi fitted a common pattern, treated above ( Figure 7 View FIGURE 7 ) and referred to as ‘Nairobi typical’ except for one caterpillar (89/35; Figure 8 View FIGURE 8 ) which was sufficiently different to suggest it was some other species, but which produced an adult male indistinguishable from others reared at Nairobi. It was collected as a penultimate instar premoult caterpillar, Kyuna Estate, Nairobi, 12 May 1989. The penultimate instar resembled a very heavily marked fourth instar of the typical form: 2.4 x 2.8mm wide x high; ground colour light brown; epicranial suture and clypeal sutures heavily black; heavy dark line adjacent to epicranial suture and contiguous with marking over epicranial suture except portion nearest vertex, widening ventrally to the top of the adfrontals and continuing to level of bottom of the frons; apical portion of adfrontals black; frons brown; dark lateral band from vertex to stemmata, widening towards stemmata; posterior margin narrowly dark. The final instar caterpillar measured 30mm when newly moulted and grew to 50mm shortly before it pupated. Head similar in size (c. 3.0 x 4.0mm wide x high), shape and orientation to Z. dysmephila Nairobi typical, but with strikingly different black and white markings; a broad white stripe, narrowed at each end, running down each side of the head; from the vertex, a short, narrow white line runs adjacent to the epicranial suture, about ¼ the distance to the adfrontals; adfrontals, except at apex and labrum white; adjacent to the dark posterior margin, there is a ventrolateral white area. By the time the caterpillar is full-grown, the white markings turn very pale brown, in this regard, resembling Nairobi typical Z. dysmephila . T1 white, with a narrow black pronotum which extends laterally to just short of the orange-brown T1 spiracle. Body translucent yellowish green; a 0.5mm wide subdorsal white stripe, yellowish T2–A2, and containing the yellow gonads at A5; spiracles conspicuous orangebrown; all legs concolorous. The pupa was not distinguished from that of Z. dysmephila Nairobi typical.
In colouring this caterpillar is closest to the caterpillars found in Kakamega Forest, Kenya (below), sharing the pale ground colour of collection 91/23 (male reared), but with markings very similar to those of Z. quaternata from Gambia ( Figure 16 View FIGURE 16 ). The genitalia seem to be the same as those of other Nairobi specimens, so the caterpillar may represent an extreme variation or an aberration of Nairobi typical.
Caterpillar, Kakamega Forest , Kenya. MJWC has reared a male and two females of what appeared to be Z. dysmephila from Kakamega Forest, western Kenya, the adults being indistinguishable in appearance from those reared in Nairobi. However, reviewing reared material for this paper in discussion with T. B. Larsen, MJWC realised that the single male reared from Kakamega (91/23) is not the same as that reared from Nairobi, based on clear differences in the male genitalia, which TBL already suspected from other parts of Africa. One individual (90/76A) was reared from an ovum and had 5 instars, matching what was found at Nairobi, and so treated as typical here.
Male 91/23 was collected 31 Mar 1991 as an instar 3 caterpillar on the same P. reclinata palm in northern Kakamega Forest as earlier collections of Z. dysmephila and of Caenides dacela (Hewitson) below. A detailed description was not prepared (at that time it was thought to be within the range of variation of Z. dysmephila ), but fortunately the last two instars were photographed ( Figure 9 View FIGURE 9 ). The head of the instar n-2 caterpillar measured 1.6 x 1.8mm wide x high; brown with the epicranial suture darker. The head of the fourth instar caterpillar measured 2.2 x 2.7mm wide x high; ground colour dark brown; a diffusely edged brown patch from near vertex to in front of stemmata; labrum brown. The head of the final instar caterpillar measured 3.1 x 3.8mm wide x high; colour almost white, with black markings: from posterior margin of vertex to stemmata, parallel to the epicranial suture, a spot on epicranium adjacent to adfrontals at level of mid frons, epicranial, adfrontal and clypeal sutures. Both instars have T1 white with a narrow black pronotum on the posterior margin. The spiracles are strikingly orange-brown.
Female 90/76A, collected 20 Jul 1990 as an ovum on the same P. reclinata was more carefully documented. The ovum was laid between the apical and adjacent leaflets, right at the base. It was 1.6 x 1.0mm diameter x height, with very fine ribs, the top flattened and dark. There were five instars, the head capsules for instars 2–5 measuring 1.2 x 1.4mm, 1.7 x 1.9mm, 2.3 x 2.8mm, and 3.2 x 4.0mm respectively (that for instar 1 was lost). Instar 1 had a black head, yellow-green body with anal segments tinted orange. Instar 2 was similar although the head was very dark brown; it measured 8mm when newly moulted. Instar 3 ( Figure 10.1 View FIGURE 10 ) measured 12mm when newly moulted; head dark brown; epicranial and clypeal sutures black; the middle of each epicranium slightly paler; black pronotum now evident; body translucent light green, white subdorsal line. Instar 4 ( Figure 10.2–4 View FIGURE 10 ) measured 25mm newly moulted; head brown; epicranial and clypeal sutures very dark; heavy dark line adjacent to epicranial suture, widening and becoming more diffuse adjacent to adfrontals, extending to level of near bottom of frons; upper part of adfrontals dark; a broad, dark lateral band from vertex to stemmata, wider ventrally; narrow band of ground colour before dark posterior margin. T1 with narrow black pronotum; body translucent dark green with white subdorsal line. This fourth instar individual resembles that of the male treated above, except the pale markings on the head are stronger. The final, fifth instar was not described or photographed; it resembles that described above for the male ( Figure 9.2 View FIGURE 9 ). Under Nairobi conditions, this individual took 9, 7, 7, 11, 19 and 23 days for the five instars and pupa.
There are differences between these early stages and those documented above as Nairobi typical, even given the level of variation observed in Nairobi typical and the small number of observations from Kakamega. Thus, in the final instar, from Kakamega Forest, the paler ground colour of the head, lateral band from vertex to stemmata, and more conspicuous orange-brown spiracles may be significant.
Caterpillar, Kenya Coast. MJWC collected Z. dysmephila at the Kenya Coast (Bamburi, Malindi) on P. reclinata , and it probably occurs throughout the developed part of the coast where ornamental P. reclinata are planted. A female and crippled male were reared from one collection made at Bamburi (90/58). Unfortunately, at the time they were thought to be the same as the caterpillars from Nairobi and so were neither photographed nor described. The following is based on an examination of the associated head capsules of the reared male and compared with all material collected at the coast. The penultimate instar head measured 1.9 x 2.1mm wide x high (n=3), rather smaller than in the same instar for Nairobi typical (2.4 x 2.8mm). However the final instar was almost the same: 3.2 x 4.1mm (n=4) compared to 3.3 x 4.2mm. Penultimate instar (90/58B): head ground colour pale brown; epicranial and clypeal sutures dark; slightly darker brown line adjacent to epicranial suture as far as adfrontals; no spot against adfrontals; laterally and around stemmata concolorous; posterior margin narrowly dark; the reared female (90/58A) was similar and another individual from Kikambala, Kilifi District (87/39), differed only in that the posterior margin was not dark. Final instar (90/58B): head ground colour pale brown; epicranial and clypeal sutures dark; dark line adjacent to epicranial suture to level with the top of the frons; dark spot on epicranium against adfrontal at about the middle of the adfrontal; patch around stemmata dark; posterior margin narrowly dark; the reared female (90/58A) was similar; in two individuals from Kikambala (87/39) the posterior margin was narrowly dark, and one of the two had a diffuse brown area laterally on the head. Thus, the caterpillars from the coast were at the pale extreme of variation observed in Nairobi typical Z. dysmephila .
Caterpillar, Rwanda. A cast final instar caterpillar skin, separate head capsules from the two previous instars and ova were collected at Butare, Rwanda on an ornamental Phoenix sp. The head of the final instar caterpillar differed from others shown here; it measured 3.0 x 4.2mm wide x high and was light brown with blackish markings: a line from vertex laterally to stemmata, widening from about half way and dividing into two basally to cover the anterior and posterior stemmata; epicranial suture dark; a strong line from just below vertex (where narrowest), parallel to epicranial suture and on epicranium adjacent on adfrontals (widest at top of adfrontals); upper half frons dark; adfrontals and posterior margin of head not dark; pronotum narrow, black; a light covering of white waxy powder; no associated pupa.
A single penultimate instar head capsule measured 2.6 x 3.1mm wide x high; light brown with heavy dark markings: posterior margin narrowly dark; epicranial and clypeal sutures dark, but adfrontal sutures not; dark line from vertex laterally to half way down head; black line parallel to epicranial suture from a little below vertex (where narrowest) to level with top of adfrontals; dark line through anterior stemmata. Three n-2 instar head capsules measured 1.8 x 2.1mm wide x high and were variable in markings. One was brown with extensive dark markings: posterior margin broadly dark to edge of face; epicranial and clypeal sutures dark; a diffuse dark area parallel to epicranial suture and on epicranium adjacent to adfrontals, from below vertex to level of mid frons, then running laterally to stemmata where it joins the dark posterior border; frons darker brown than brown of face. The other two were dark, with slightly paler areas on the upper epicranium and from the lower half of the adfrontals to the ventral stemmata, similar to the third instar caterpillar from western Kenya shown in Figure 10.1 View FIGURE 10 .
Caterpillar, Mafwemiru, Rubeho Mountains, Tanzania. A 2013 collection by TCEC on Phoenix reclinata in the Rubeho Mountains produced Z. dysmephila caterpillars with a pale brown head, no lateral line, a strong black line parallel to the dark epicranial suture, a spot around the stemmata, and individuals with and without a spot adjacent to the lower adfrontals ( Figure 11 View FIGURE 11 ). These caterpillars resemble those documented above from Nairobi, but not elsewhere.
Caterpillar, Mufindi, Southern Highlands of Tanzania. A caterpillar from Mufindi ( Figure 12 View FIGURE 12 ) has the pale brown head of Nairobi ( Figure 7 View FIGURE 7 ) and southern African populations. The heavy dark line adjacent to the epicranial suture is separate from a lower black spot adjacent to the adfrontal suture, resembling individuals from Kenya (all three populations, Figures 7 View FIGURE 7 , 9 View FIGURE 9 , 10 View FIGURE 10 ), central Tanzania ( Figure 12 View FIGURE 12 ) and north-east Zambia ( Figure 13 View FIGURE 13 ), but not elsewhere. There is a heavy black band from the dorsal posterior margin to the stemmata, widening basally, resembling western Kenya specimens ( Figure 9, 9 View FIGURE 9 ), and that documented from Zambia below, but not those from Central Tanzania ( Figure 11 View FIGURE 11 ), Nairobi ( Figure 7 View FIGURE 7 ) or the Kenya Coast.
Caterpillar, north-east Zambia. Caterpillars recently collected by TCEC in north-east Zambia have the head pale brown, strongly marked with black, similar to the most heavily marked Nairobi examples ( Figure 7.2 View FIGURE 7 ), with a dark lateral line, although not as strong as that of the individual from Mufindi, Tanzania; the body had yellow subdorsal lines rather than white ( Figure 13 View FIGURE 13 ). It appears to be almost identical otherwise to the caterpillar from Mufindi, Tanzania.
Pupa. The pupa (based on Nairobi typical material, Figure 14 View FIGURE 14 ) is 28mm long; head, appendages and dorsal areas dark brown; abdomen pale brown ventrally; becoming darker during development; surface shiny under a light covering of white waxy powder; rounded, with the frons and eyes slightly protuberant; thorax and eyes (except vertical bar down middle) with light covering of reddish brown, erect setae; the proboscis does not protrude beyond the wing cases. The inside of the shelter is heavily covered with white waxy powder, but the pupa itself only lightly so. The pupae from other populations did not appear to differ. The pupal stage lasts about 18 days in Nairobi (range 17–21, n=3), compared to 18–25 days at 2300m in Eritrea ( Jannone 1948).
Natural enemies. South Africa. Dickson & Kroon (1978) record?cynipid and chalcid egg parasitoids, Pimpla sp. (ichneumonid third instar caterpillar parasitoid), tachinid larval-pupal parasitoids Nemorilla cruciata (Wiedemann) , Erycia sp. and Theocarcelia incedens (Rondani) (?synonym of T. acutangula (Macquart)) , and chalcid pupal parasitoids. They also note that insectivorous birds take a heavy toll of caterpillars.
Nairobi , Kenya. The ova are parasitized by gregarious trichogrammatid egg parasitoids; for example one produced 6 tiny, brown adult wasps 18 days after it was collected (Nairobi, 90/20). On two occasions, MJWC found dead caterpillars in their shelters in Nairobi, one associated with four pupae (90/122) which produced one male and three females of an? Elasmus sp. (Eulophidae) and another (fourth instar, 90/96) with nine naked black pupae of a euplectrine, which emerged 3–4 days later, yielding one male and seven female adults (and one escape) .
Kakamega, Kenya. Twice MJWC found large caterpillars in North Kakamega Forest parasitized by a small gregarious tachinid fly. A fourth instar caterpillar produced 3 puparia which failed to emerge, and a fifth instar caterpillar produced 13 small puparia, but only one crippled adult fly emerged. It appears to be the same species, Ceranthia sp. , which has also been reared from Hesperiinae Pelopidas mathias (Fabricius) ( Cock & Congdon 2012) and Chondrolepis leggei (Heron) (MJWC unpublished) in western Kenya.
Kenya Coast. At the Kenya coast the ova are attacked by a dark egg parasitoid, larger than those mentioned above. An ovum collected on a small, but partially defoliated Phoenix reclinata near Malindi, 22 Sep 1989 ( Figure 3.1 View FIGURE 3 ) produced a single such parasitoid on 5 Oct ( MJWC 89 /63) .
Rwanda. The material collected at Rwanda (90/202) revealed a high level of egg parasitism. In the batch of 24 ova, three were completely missing having probably become detached from the substrate, three had eclosed normally, 14 had parasitoid emergence holes and 4 were dead, perhaps due to parasitism. Two individually laid ova produced two different species of solitary egg parasitoid 21 and 22 days after collection. One small leaf shelter typical of Z. dysmephila contained three black euplectrine pupae, one of which emerged seven days later.
Discussion. We have shown geographical and local variation in the caterpillars of Z. dysmephila , some of which correlates with unanticipated variation in male genitalia, where this has been investigated. More rearing is needed to assess the extent of variation at most sites, as it is not clear whether this variation reflects a variable species throughout its range, or is a function of geography. The material from Nairobi suggests a discrete, constant form of the male genitalia, but there is not enough material in series from other areas to assess the observed differences reported here. Zophopetes dysmephila is a complex species, or a species-complex, and more rearing, male and female genitalia examination and barcoding will be needed to work out the full picture. This needs to be followed up with further study.













| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Hesperiinae |
|
Genus |
Zophopetes dysmephila Trimen, 1868
| Cock, Matthew J. W., Congdon, Colin E. & Collins, Steve C. 2014 |
Z. dysmephila
| Trimen 1868 |
Z. dysmephila
| Trimen 1868 |