
Cirratulus balaenophilus, Taboada, Sergi, Doner, Stacy, Blake, James A. & Avila, Conxita, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.211639 |
|
DOI |
https://doi.org/10.5281/zenodo.5670028 |
|
persistent identifier |
https://treatment.plazi.org/id/62652661-DA2A-FFA4-FF14-7F0CFC2A4CB6 |
|
treatment provided by |
Plazi |
|
scientific name |
Cirratulus balaenophilus |
| status |
sp. nov. |
Cirratulus balaenophilus View in CoL sp. nov.
Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4
Material examined. Port Foster, Deception Island, South Shetland Islands, Antarctica; 62º59.33' S; 60º33.45' W, 21 m, associated with a minke whale ( Balaenoptera acutorostrata ) caudal vertebra. Collected by S. Taboada, J. Cristobo, and C. Avila, January 25, 2010. Holotype (CRBA-9616) and four paratypes (CRBA-9617; CRBA-9618; CRBA-9619; CRBA-9620) complete and well preserved. Additional material: seven specimens preserved in 10% formalin and transferred to 70% ETOH, three used for SEM; 10 specimens preserved in 95% ETOH.
Description. A moderate-sized species; holotype complete, 14 mm long, 0.6 mm wide across both thorax and abdomen, for about 100 chaetigers ( Fig. 2 View FIGURE 2 A). Largest paratype complete, 7.2 mm long and 0.5 mm wide across both thorax and abdomen, for about 69 chaetigers ( Fig. 2 View FIGURE 2 B). Remaining paratypes all complete juveniles, 2.6–3.6 mm long, 0.2–0.3 mm wide across both thorax and abdomen, with 40–45 chaetigers. Thoracic region not expanded, composed of 11–15 crowded chaetigers, lacking dorsal and ventral grooves. Juveniles with about six crowded thoracic chaetigers. Cross-section of thoracic region rounded both dorsal and ventrally. In life, body light yellow-orange with conspicuous dorsal red blood vessel anteriorly; tentacles pale yellow ( Fig. 2 View FIGURE 2 B). After preservation specimens opaque white, lacking all pigmentation; dorsal blood vessel remains visible ( Fig. 2 View FIGURE 2 A).
Prostomium as wide as long, anteriorly rounded; eyes absent; peristomium longer than wide with three prominent annulations viewed both dorsal and ventrally ( Fig. 3 View FIGURE 3 A–B). Pair of circular nuchal organs located posterior and lateral to the mouth, lacking pigmentation ( Fig. 3 View FIGURE 3 B, 4C–D). Dorsal tentacles arising from chaetiger 1, arranged in two groups, each with up to four separate tentacles forming an arc ( Fig. 3 View FIGURE 3 A); first branchiae arising just dorsal to notochaete on chaetiger 1 ( Fig. 3 View FIGURE 3 A–B, 4A–B). Second pair of branchiae dorsal to notochaetae on chaetiger 2, in line with first pair; subsequent branchiae close to notochaeta throughout. Branchiae thick in anteriormost chaetigers decreasing in thickness posteriorly.
Noto- and neurochaetae arise close together throughout, shifted ventrally in posterior segments. Each podial lobe bearing long simple capillaries lacking serrations; 6–9 capillaries per fascicle anteriorly arranged in two rows; decreasing in posterior region to 2–3 capillaries in single row. Noto- and neurochaetae similar in length throughout, appearing smooth under the light microscope, with splayed fibrils observed in SEM pictures ( Fig. 4 View FIGURE 4 E).
Thoracic segments with well-developed parapodia ( Fig. 3 View FIGURE 3 B, 4B); reduced to low tori posteriorly. Thoracicabdominal transition indistinct. Abdominal segments slightly less crowded, never moniliform. Posterior end weakly tapered, dorsally rounded and ventrally flattened ( Fig 2 View FIGURE 2 C). Pygidium ventral, simple rounded lobe above terminal anus ( Fig. 3 View FIGURE 3 C).
Methyl Green staining. Methyl Green staining observed only in adults; conspicuous ventral staining of chaetigers 10–20. Remaining chaetigers of first half of body with thin band on posterior half of segment, incomplete ventrally ( Fig. 2 View FIGURE 2 C–D).
DNA analysis. The cytochrome c oxidase subunit I (COI) sequence was obtained from the sequencing vouchers (MCZ 106471) and is published on GenBank ( JQ048545 View Materials ). The 28S sequence was also attempted without success.
Distribution. Cirratulus balaenophilus sp. nov. is known only from Port Foster, Deception Island, in the South Shetland Islands, Antarctica.
Habitat and ecology. As oxygen was depleted from the water in which the bone was kept, the yellowish tentacles of C. balaenophilus sp. nov. were noticed in the galleries of the trabecular bone tissue ( Fig. 2 View FIGURE 2 E). These organisms apparently lived inside these natural cavities and did not appear to bore into the bone. This species may live only in fresh whale bones since it did not occur in an older vertebra with no smell of sulphide and with no apparent remaining organic matter that was collected from a similar depth in a nearby area ( Fig. 1 View FIGURE 1. A C; Sta. 2). It is important to point out that this old bone, presumably dating back to the whaling factory that operated in Whalers Bay in the early 20th century, hosted other polychaete species that were also found in the experimental fresh bone (ST & CA unpublished data). Thus, we may hypothesize that C. balaenophilus sp. nov. relies on the food supply that the fresh bone provides. No information about reproduction can be inferred from the specimens collected due to lack of ovigerous specimens.
Tentacles
Area Prostomium Num. Capillaries Color after
Species Eyes location Spines Data source
Depth range shape Chaetigers remarks preservation (num. pairs)
Remarks. The absence of spines, the number and arrangement of the dorsal tentacles, and the lack of eyes characterize Cirratulus balaenophilus sp. nov. Of the six species of Cirratulus occurring in the Southern Ocean and adjacent waters, only C. nuchalis ( Ehlers, 1907) and C. parafiliformis Hartmann-Schröder & Rosenfeldt, 1989 are also reported to lack spines ( Table 1 View TABLE 1 ). C. nuchalis was originally described from the harbor of Auckland, New Zealand, and differs from C. balaenophilus sp. nov. in the prostomial shape (rounded rather than triangular), color in life (purple as opposed to light yellow-orange), the number of chaetigers in adults (375 versus 40–100), and the very long branchiae that is its most characteristic feature.
On the other hand, C. parafiliformis was originally described from an area near Deception Island (Admiralty Bay, King George Island), although from deeper waters (133–458 m). That species, as described, differs from C. balaenophilus sp. nov. in prostomial shape (bluntly conical versus triangular), the number of dorsal tentacles (5–6 pairs rather than 3–4), in having capillary chaetae of different sizes, as well as in the color after preservation (dark brown-purple instead of white) ( Table 1 View TABLE 1 ). As part of a separate study, one of us (JAB) examined 43 paratypes (ZMH-19622) of C. parafiliformis deposited at the Zoologisches Museum der Universität Hamburg and can confirm that C. parafiliformis is indeed a different species than C. balaenophilus sp. nov. The body of C. parafiliformis is relatively robust; the thoracic region consists of numerous crowded segments; abdominal segments are generally narrow and more elongate, but not moniliform. The body is flesh-colored in alcohol; some specimens have a dusky black pigmentation on the dorsum, sides, and ventrum of the first 5–10 chaetigers. The prostomium is triangular in outline, tapering to a narrow anterior end; eyes are absent. There are three peristomial annulations as in C. balaenophilus sp. nov., but the entire head region is compressed and thicker. Dorsal tentacles occur anterior to chaetiger 1 and number 5–7 in each of two groups. All chaetae are capillaries and are arranged in spreading fascicles with the chaetae of anterior segments long and silky. The pygidium is a simple ring surrounding the anus without appendages. Methyl Green imparts a distinctive “mask” to the head region, with each of the three peristomial annulations staining heavily with a clear dorsal unstained area between the first and second annulations and smaller clear areas laterally; the tip of the prostomium does not stain; anterior chaetigers retain stain intersegmentally mainly on the lateral and ventral surfaces.
Regarding the presence or absence of eyes, most species of Cirratulus occurring in shallow waters tend to have eyes (e.g., C. cirratus sensu Hartman, 1966 and C. gilchristi Day, 1961 ). However, species such as C. balaenophilus sp. nov., C. nuchalis , and C. patagonicus ( Kinberg, 1866) , also shallow waters dwellers, lack eyes.
Etymology. Cirratulus balaenophilus sp. nov. is named after its original collection from a whale bone as well as from having been collected from the area of Whalers Bay in Deception Island. Balaenophilus is a Latin-Greek compound word meaning “liking whales” (balaena = whale, philus = like).


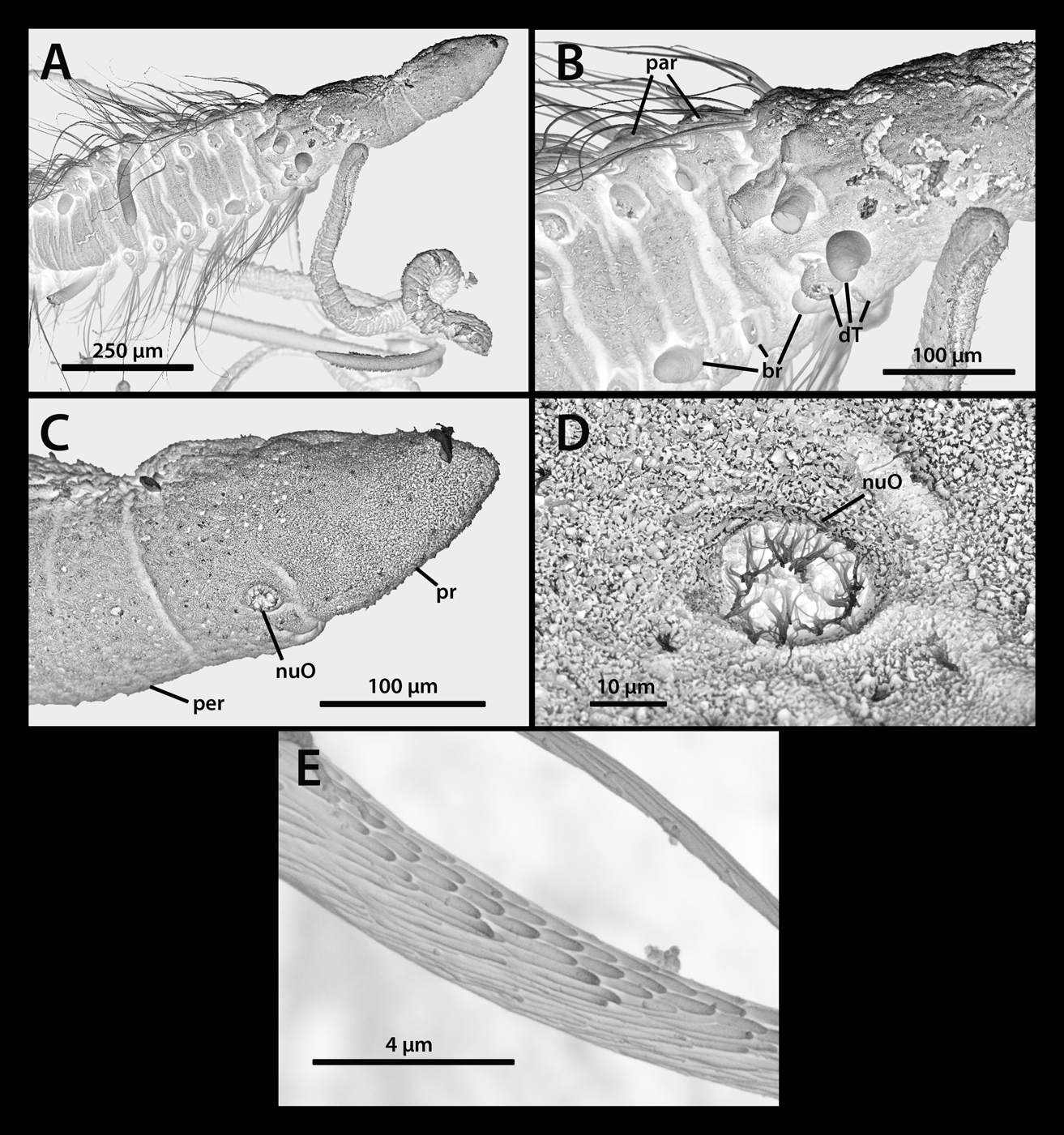

TABLE 1. Comparative list of characters for Cirratulus species found in the Southern Ocean and adjacent waters.
| Cirratulus cirratus 1 (O. F. Müller, 1776) | Cosmopolitan. Intertidal to 845 m | bluntly conical | yes | 93–130 | chaetiger 1 (7 p.) | dorsal longer than ventral | yes | Hartman 1966, 1967; deep purple-pale Carrasco 1977 |
|---|---|---|---|---|---|---|---|---|
| Cirratulus concinnus Ehlers, 1908 Cirratulus gilchristi Day, 1961 | Agulhas Bank (South Africa). 117 m Saldanha Bay (South Africa). 8–12 m | conical, ventrally flattened broadly rounded | no yes | 77–80 98 | perist.- chaetiger 1 (1–2 p.) perist.- chaetiger 1 (8 p.) | long throughout nothing remarkable | yes yes | — Ehlers 1908 pale yellow Day 1961 |
| Cirratulus nuchalis (Ehlers, 1907) | Auckland (New Zealand). Shallow waters | rounded | no | 375 | — | shiny | no | red-brown Ehlers 1907 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |