
Glomeridesmidae Latzel, 1884
|
publication ID |
https://doi.org/ 10.5281/zenodo.211959 |
|
DOI |
https://doi.org/10.5281/zenodo.6179706 |
|
persistent identifier |
https://treatment.plazi.org/id/5571F65F-9C13-DA25-05D8-8AA4FC1A6D34 |
|
treatment provided by |
Plazi |
|
scientific name |
Glomeridesmidae Latzel, 1884 |
| status |
|
Family Glomeridesmidae Latzel, 1884 View in CoL
Zephroniodesmidae Cook, 1895: 2
Comment: A monogeneric family ( Jeekel 2003, Shear 2011).
Characters: Females with 36 pair of legs, 20 tergites plus anal shield; males with 35 leg pairs, 19 tergites plus anal shield (see Hoffman 1982, Enghoff et al. 1993). Males with 20 tergites and 37 leg pairs are supposed to exist but have not yet been discovered ( Carl 1942).
Head without ocelli, but with large, circular organ of Tömösváry ( Fig. 3 View FIGURE 3. G. s p e l a e u s n D). Genae (head sclerites below the antennae) absent. Antennae long, slender, with seven joints and an apical disc carrying four long, slender apical cones ( Fig. 3 View FIGURE 3. G. s p e l a e u s n F). The mandible seems to consist of a sutureless fused cardo and stipites ( Fig. 3 View FIGURE 3. G. s p e l a e u s n C), confirming earlier observations ( Carl 1942).
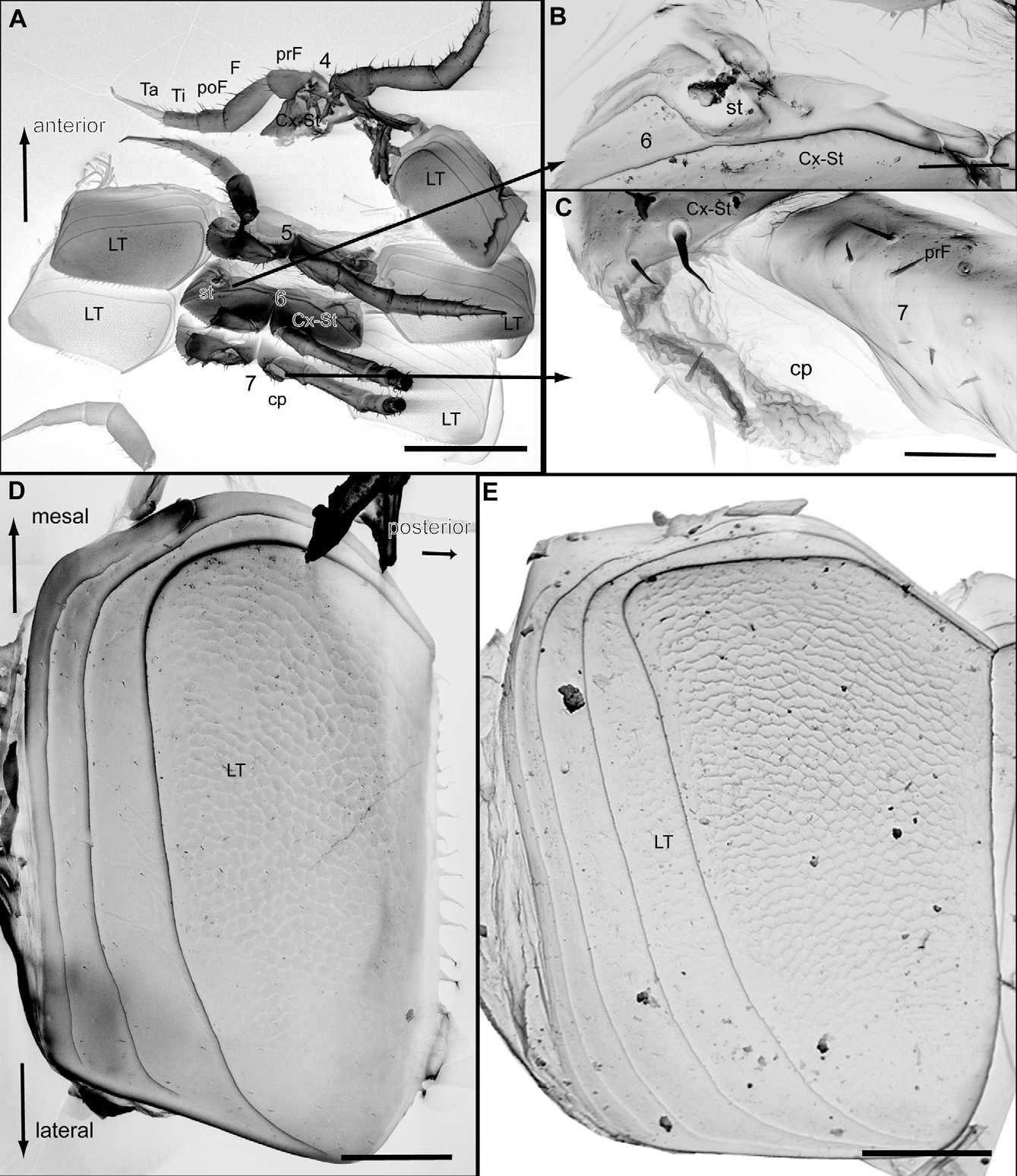
Each body segment composed of tergite forming a semicircle, one large laterotergite ('pleurite') on each side, and two coxosternites with stigma and tracheal apodeme ( Fig. 3 View FIGURE 3. G. s p e l a e u s n A, 4A, B). As in earlier papers on the phylogeny of Pentazonia (Wesener & VandenSpiegel 2009), we follow here Snodgrass’ view of the so-called pleurites as laterotergites ( Snodgrass 1958). Tergites generally soft, weakly calcified, not divided into pro-, meso- and metazona, but with numerous (5–10) longitudinal striae ( Fig. 6 View FIGURE 6. G A, B, E). Lateral tips of last 3–5 tergites always(?) projecting posteriorly, overlapping following tergites with a sharp-edged tip ( Fig. 6 View FIGURE 6. G A, B). Ozopores for secretion of defense fluid absent.
Development hemianamorphic, leg and tergite development apparently decoupled ( Enghoff et al. 1993). Last leg pair of males (either 33rd, 34th or 35th) modified to telopods ( Fig. 8 View FIGURE 8. G. s p e l a e u s n A, B, 9A, B). Last leg in females, as well as second last in males, characteristically modified, with undivided large sternal plate ( Fig. 7 View FIGURE 7. G. s p e l a e u s n A), tarsus posteriorly protruding above anal shield ( Fig. 7 View FIGURE 7. G. s p e l a e u s n B). Starting at the mid-body each odd-numbered coxa (e.g. 11, 13, 15) with eversible coxal pouch ( Fig. 4 View FIGURE 4. G. s p e l a e u s n A, C).
Gonopores in both sexes located on posterior side of coxa 2. Females with long ovipositors (misinterpreted as 'penes' by earlier authors) forming long (up to half of body length), eversible, membranous tubes ( Fig. 3 View FIGURE 3. G. s p e l a e u s n B). Male gonopore consisting of two small sclerotized plates located mesally on the coxa ( Fig. 5 View FIGURE 5. G A, B).
Telopods (only known from 7 species) divided into a syncoxite with inner horns ( Fig. 8 View FIGURE 8. G. s p e l a e u s n A) and a 4-jointed telopod. Joint 2 at apico-mesal margin with a species-specific (?) membranous field, joint 3 elongated into immovable finger located laterally (see Wesener & VandenSpiegel 2009: fig. 37) or posteriorly ( Fig. 9 View FIGURE 9. G. s p e l a e u s n A) of joint 4, which forms the movable finger and carries a prominent apical spine ( Fig. 9 View FIGURE 9. G. s p e l a e u s n A, B, D).
The complete inner morphology remains unstudied (see discussion below).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.