
Notoalona pseudomacronyx Van Damme, Maiphae & Sa-Ardrit, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4615.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:290502EB-A15A-4699-B063-BDB7084A90C1 |
|
persistent identifier |
https://treatment.plazi.org/id/4B4BC250-FF80-3762-FF0D-0358088DFB8E |
|
treatment provided by |
Plazi |
|
scientific name |
Notoalona pseudomacronyx Van Damme, Maiphae & Sa-Ardrit, 2013 |
| status |
|
Notoalona pseudomacronyx Van Damme, Maiphae & Sa-Ardrit, 2013
( Figs. 6–10 View FIGURE 6 View FIGURE 8 View FIGURE 10 )
Notoalona pseudomacronyx in Van Damme, Maiphae & Sa-Ardrit 2013a: p. 176 –182, figs. 1–3.
Alona globulosa Daday, 1898 in Kořínek 1984: p. 62 –63, pl. XL, figs. 1–10.
Alona globulosa Daday, 1898 in Rey & Saint-Jean 1968: p. 113 , figs. 28a–c.
Indialona globulosa ( Daday, 1898) in Fernando 1974 : figs. 116–118.
? Indialona globulosa insulcata (Stingelin) in Dumont et al. 1981: p. 165 (without figures).
Etymology. The species epithet pseudomacronyx refers to similarity of postabdomen of this species with postabdomen of Celsinotum macronyx ( Daday, 1898) (see Sinev & Kotov 2012; Van Damme et al. 2013a).
Type locality. Natam swamp, Trang province, western coast of South Thailand ( Van Damme et al. 2013a).
Type material.
Holotype. An adult parthenogenetic female in a sealed 70% ethanol, PSUZC-PK1001-01 .
Paratypes: 2 adult parthenogenetic females, PSUZC-PK 1001-03 and PSUZC-PK 1001-04 ; 5 adult parthenogenetic females in a tube with 70% ethanol, PSUZC-PK 1001-02 ( Van Damme et al. 2013a) .
Material studied from Africa. GoogleMaps
Ethiopia: 2 parthenogenetic females from a temporary pool on the road between Hamusit & Worota (N 11.82511°, E 37.60931°), coll. 24.09.2015 by W. Zelalem, ANN 2016 -002; 1 parthenogenetic female from a temporary pool on the road between Hamusit & Worota (N 11.82511°, E 37.60931°), coll. 24.09.2015 by W. Zelalem, ANN M-0113 GoogleMaps .
Kenya: 3 parthenogenetic females from unknown locality, coll. by R. Smolak, AAK 2016 -057 .
Sudan: 8 parthenogenetic females from River Sobat , right affluent of the White Nile, coll. 26.12.1963 by A.V. Monakov, NMK 1533 View Materials ; 5 parthenogenetic females from the White Nile near Gerer, coll. 11.12.1963 by A.V. Monakov, NMK 1534 View Materials ; 8 parthenogenetic females from the White Nile (near Aljab ?), coll. 27.01.1964 by A.V. Monakov, NMK 1536 View Materials .
Madagascar: 2 parthenogenetic females from Lac Anatanavo, Montagne d’Ambre National park, coll. 19.05.2009 by R. Schabetsberger, AAK 2010 -010.
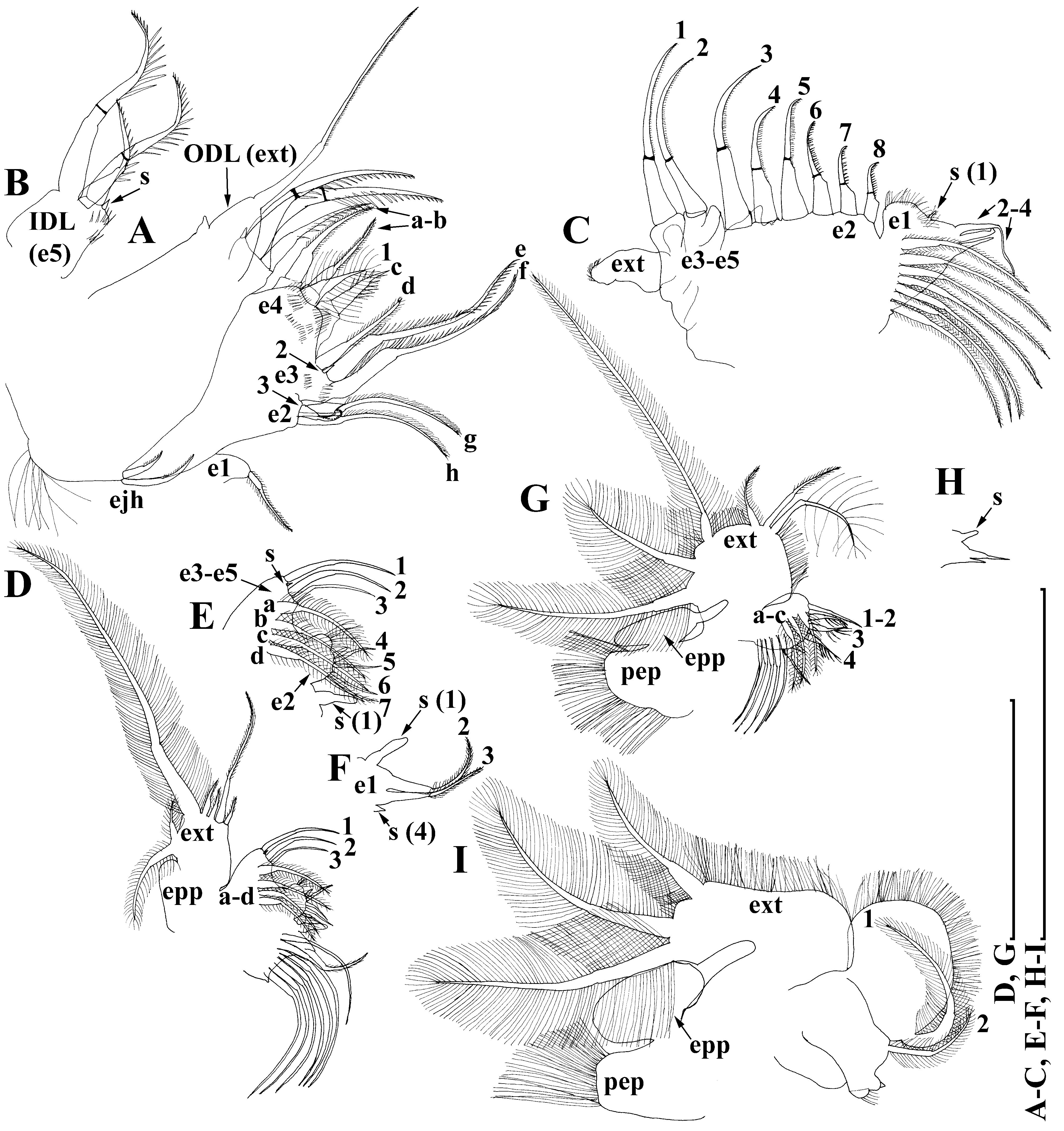
Description of African populations. Parthenogenetic female. General. Body subglobular, without expressed dorsal keel, maximum height anterior to body middle (body height/length ratio about 0.85 for all investigated specimens) ( Figs. 6A View FIGURE 6 , 7A, F, 8A, 9A) basically similar with N. globulosa . Shape, proportions and armature of head ( Figs. 6 View FIGURE 6 A–C, 7A–B, F, 8A–D, 9A), labrum ( Figs. 6D View FIGURE 6 , 7H–G, 8A, 9A–B), valve ( Figs. 6 View FIGURE 6 E–I, 7A, C, F, 8A, 9A), thorax ( Fig. 6A View FIGURE 6 ), abdomen ( Fig. 6A View FIGURE 6 ) and postabdomen (6J, 9C–D) are identical to previously described species.
Postabdominal claw long and narrow (subequal in length to anal margin of postabdomen), and moderately curved ( Figs. 6 View FIGURE 6 J–L, 7I, 8E, 9C–D). Armature of outer and inner sides of claw ( Figs. 6 View FIGURE 6 J–L, 7D–E, I–J, 8E, F, 9C–E) is similar with N. globulosa as well. However, the basal spine in N. pseudomacronyx is relatively long, 2 times longer than the diameter of postabdominal claw at its base ( Figs. 6 View FIGURE 6 K–L, 7I, 8E, F, 9E). A bunch of fine setules present F IGURE 7. Notoalona pseudomacronyx Van Damme, Maiphae & Sa-Ardrit, 2013 , two parthenogenetic females from the White Nile near Gerer, Sudan, coll. 11.12.1963 by A.V. Monakov. A, F, Adult parthenogenetic female, lateral view. B, Head. C, Armature of ventral portion of valve. D–E, I–J, Fragments of postabdominal claws. G, Labral keel. H, Branches of antenna II. Scale bars 0.1 mm for A, F, 0.05 mm for B, 0.02 mm for G–I, 0.01 mm for C–D, 0.005 mm for E, J.
Ephippial female, male. No ephippial females and males were found in our material. Also, ephippial females and males were not found by Van Damme et al. (2013a).
Size. In our material, adult parthenogenetic females up to 0.42 mm in length and 0.38 mm in height. Values of length published by Van Damme et al. (2013a) are less, mostly 0.285 mm (never exceed 0.3 mm), but, probably, only the restricted number of individuals were measured.
Variability. No significant variability was found between all investigated African populations. Some minor variability concerns lengths of setules in the bunch near postabdominal basal spine. The same observation was already made by Rey & Saint-Jean (1968: fig. 28b–c, very short setules in the bunch near postabdominal basal spine) and Kořínek (1984: pl. XL, figs. 6–8, relatively long setules).
Differential diagnosis. Postabdominal basal spine two times longer than the diameter of claw at its base (see Table 1 View TABLE 1 ), a group of fine setules is present near basal spine (longest setules reach almost half of basal spine).
Taxonomic remarks. In Africa we found only populations with long postabdominal basal spines (this spine is 2 times longer than diameter of postabdominal claw at its base) (see Table 1 View TABLE 1 ). Thus, we may confidently identify them as N. pseudomacronyx in contrast to Rajapaksa & Fernando (1987a) who considered them as N. globulosa globulosa . At the same time, we found some variability in the length of setules in the bunch near postabdominal basal spine. Therefore, in our opinion, this feature could not be used for description of African populations as a separate species in contrast to Van Damme et al. (2013a) and Neretina et al. (2017). Dubiousness of this feature was reflected even in the text and figures published by these authors (in figure 1l–m for Thai N. pseudomacronyx setules reach a half of postabdominal basal spine length; in figure 12 for Thai N. pseudomacronyx they are more short (up to 1/4 of postabdominal basal spine length), while in Notoalona from Africa, labeled as “sp.” these setules are also long). More likely, that a degree of setules development is correlated with length of postabdominal basal spine.
Distribution and ecology. N. pseudomacronyx is known from several African localities ( Rey & Saint-Jean 1968: figs. 28a, b, c; Kořínek 1984: pl. XL, figs. 1–10; Rajapaksa & Fernando 1987a: figs. 53–55, 57–59, discus- sion for fig. 56 see below; Schabetsberger et al. 2013 —studied material was partially re-examined in our work), but a finding of this taxon in Madagascar allows us to presume, that, in fact, it has a wide distribution range on the continent. At the same time, N. pseudomacronyx was not abundant in the samples, therefore this taxon could be considered as rare. This was also the case in the original samples examined from Southern Thailand by Van Damme et al. (2013a), where the species was found only in a sole locality despite intense sampling efforts. Also, due to its small size, N. pseudomacronyx could be confused with some other small-sized chydorids for the untrained eye (like juvenile specimens of Leberis or general Alona- like taxa) during a superficial sample analysis under a binocular stereoscopic microscope. According to our observation and literature data (e.g. Monakov 1968; Rey & Saint-Jean 1968), in Africa N. pseudomacronyx inhabits swampy areas (temporary large pools with vegetation, flooded vicinities of lakes and rivers) and they are probably associated with wet season (our observations on Ethiopian localities), but investigations on ecology of this taxon in Africa must be continued in the future. Thus, distribution range of N. pseudomacronyx is not restricted to Natam swamp in Southern Thailand in South East Asia, but also expands on Africa.
During revision on available references on Asian populations on Notoalona , we found that one record from Sri Lanka, published by Fernando (1974), in fact, belongs to N. pseudomacronyx , not to N. globulosa , due to presence of long postabdominal basal spine (see in Fernando 1974: fig. 118).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aloninae |
|
Genus |
Notoalona pseudomacronyx Van Damme, Maiphae & Sa-Ardrit, 2013
| Neretina, Anna N., Kotov, Alexey A. & Damme, Kay Van 2019 |
Notoalona pseudomacronyx
| in Van Damme, Maiphae & Sa-Ardrit 2013: 176 |
Alona globulosa Daday, 1898 in Kořínek 1984 : p. 62
| Daday, 1898 in Korinek 1984 |
Indialona globulosa insulcata (Stingelin)
| in Dumont 1981: 165 |
Indialona globulosa ( Daday, 1898 )
| in Fernando 1974 |
Alona globulosa
| Daday, 1898 in Rey & Saint-Jean 1968 |