
Tragia peltata Vellozo (1831b: 6)
|
publication ID |
https://doi.org/10.11646/phytotaxa.570.1.6 |
|
DOI |
https://doi.org/10.5281/zenodo.7258409 |
|
persistent identifier |
https://treatment.plazi.org/id/450087EC-FFC4-F620-FF3C-FB2EFD8EFC10 |
|
treatment provided by |
Plazi (2022-10-26 07:11:48, last updated 2024-11-29 00:58:39) |
|
scientific name |
Tragia peltata Vellozo (1831b: 6) |
| status |
|
Tragia peltata Vellozo (1831b: 6) View in CoL .
Lectotype (designated here):— BRAZIL. Rio de Janeiro: Paraty, Vellozo s.n. (illustration in the original manuscript of Flora Fluminensis in the Manuscripts Section of the National Library of Rio de Janeiro [cat. no.: mss1198659_010] and later published in Vellozo, Fl. Flumin. Icones 10: t. 6. 1831b). Figure 1 View FIGURE 1 .
Subshrubs, scandent, glabrescent, stems 2–3 mm diam., internodes 1.5–8.5 cm long; internodes with rare stinging trichomes and dense simple non glandular trichomes. Stipules triangular, ca. 2–3(–4) × 1–2 mm, glabrescent, with rare stinging trichomes, deciduous. Leaves alternate, spirally arranged; petiole 20(–40) × 1–1.5 mm, cylindrical, hirsute; leaf blade 8.5–14.5 × 4–7 cm, peltate, attachment distance 1–10 mm, ovate or oblong, base emarginate to retusecordate, apex acuminate, margins sparsely serrated or subentire, teeth acuminate, starting from the mid-region of the leaf towards the apex, adaxial and abaxial faces glabrescent, with sparse stinging trichomes; primary basal nerves actinodromous, secondary nerves craspedodromous or brochidodromous, 5–8 pair. Inflorescences axillary, bisexual, racemiform thyrses, usually grouped in 2–4; axis 4–6.5 cm long, with short stipitate glandular trichomes, bracts 1- flowered, basal bract 1–1.5 mm long, 3-lobed, lobes triangular, apex acute, the other bracts ca. 1 × 0.5 mm, triangular, apex acute. Staminate flowers single per node, pedicel 2–3(–4.5) mm long, hispidulous, with stinging trichomes and short stipitate glandular trichomes; sepals 3–4, ca. 1–1.2 × 0.5–0.8 mm, elliptic, apex acute, adaxial face glabrous, abaxial face with rare stinging trichomes; stamens 2–3, filaments 0.5–0.8 mm long, with central gland, anthers oblong, ca. 0.3 mm long. Pistillate flowers solitary at the base of the inflorescence; pedicel 13–20 mm long, with hispidulous, sparse stinging trichomes, and abundant subsessile glandular trichomes; sepals 6(–8), ca. 2.5 × 1.2 mm, ovate, apex acuminate, apical gland present, margins ciliate with stipitate glandular trichomes, adaxial surface glabrous, abaxial surface hispidulous, with sparse stinging and more abundant glandular trichomes; ovary 0.5–0.7 × 1 mm, sessile, 3-lobed, each lobe 2-corniculated, hispidulous, covered with stinging trichomes, styles 1.2–2 mm long, joined for approximately 3/4 of their length, stigma smooth. Fruit ca. 6 × 10–15 mm, transversely ellipsoid, 3-lobed, lobes 2- corniculate, cornicles 3–5 mm long, hispid; peduncle 25–30 mm long. Seeds ca. 4.5 × 4 mm, globose, smooth, light brown spots formed by small elliptical papillae. Figure 2 View FIGURE 2 .
Material examined:— BRAZIL. Bahia: Itamaraju , 16°59’48”S, 39°33’12”W, 5 May 1971, fl., fr., T.S. Santos 1551 (CEPEC) GoogleMaps . Espírito Santo: Linhares , 19°23’28”S, 40°04’20”W, 13 November 2010, fl., D.A. Folli 6736 (SP) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 12 April 1993, fl., fr., D.A. Folli 1864 (CVRD, K) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 09 December 1993, fl., fr., D.A. Folli 2117 (CVRD, NY, SP) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 01 December 1998, fl., fr., D.A. Folli 3306 (CVRD, SP) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 15 October 2003, fl., fr., D.A. Folli 4631 (CVRD, SP) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 26 November 2004, fl., fr., D.A. Folli 4982 (CVRD, SP) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 09 December 1993, fl., D.A. Folli s.n. (IPA) GoogleMaps ; Linhares, 19°23’28”S, 40°04’20”W, 15 October 2003, fl., fr., D.A. Folli 4632 (VALE) GoogleMaps ; Linhares 19°23’28”S, 40°04’20”W, 26 November 2004, fl., fr., D.A. Folli 4982 GoogleMaps ; Sooretama, 19°01’03”S, 40°08’43”W, 26 October 2012, fl., fr., G.O. Romão 1160 (MBML) GoogleMaps . Rio de Janeiro: Angra dos Reis, 23°03’34”S, 44°18’51”W, 2–9 September 1980, fl., D. Sucre & J. Gomes 11277 (RB); unknown location, unknown date, fl., fr., Talbot s.n. (K 001205063) GoogleMaps .
Taxonomic Considerations:— Tragia peltata was characterized in the literature as having peltate leaves, sparsely serrate, with teeth from the mid-leaf to the blade apex ( Vellozo 1831b, 1881, Müller Argoviensis 1866, 1874, Pax & Hoffmann 1919), although the morphological analyses performed here indicate previously unknown variations for this species.
The leaf is consistently peltate, but the distance between the base of the blade and the petiole insertion point is variable ( Fig. 3A, B, E View FIGURE 3 ), giving the impression that some leaves are not peltate ( Fig. 3E View FIGURE 3 ). The recognition of this character state is fundamental to the identification of T. peltata , as this character is unique among the South American species ( Pax & Hoffmann 1919). Sparsely serrated leaf margins occur only in specimens from Rio de Janeiro State ( Fig. 3 A, C View FIGURE 3 ), while the margins of specimens from Bahia and Espírito Santo are predominantly subentire ( Fig. 3 D, F View FIGURE 3 ). Additionally, the leaves of the specimens from Rio de Janeiro are membranous, while those of specimens from Bahia and Espírito Santo are chartaceous.
Tragia peltata is morphologically related to T. aurea Rusby (1907: 444) , T. giardelliae Gutiérrez de Sanguinetti & Múlgura de Romero (1986: 491) , T. hoffmanniae W.Cordeiro & M.F.Sales (in Cordeiro et al. 2020a: 841), and T. volubilis Linnaeus (1753: 980) ( Table 1 View TABLE 1 ). These species share an accrescent pedicel of the pistillate flower, which can attain the size of the axis of the inflorescence ( Cordeiro et al. 2020b). This is an important taxonomic feature that may aid in the systematics of the genus ( Cardinal-McTeague & Gillespie 2016). Despite having this characteristic in common with the abovementioned species, T. peltata can easily be recognized by its peltate leaf base ( Fig. 2 B View FIGURE 2 ) and emarginate to retuse-cordate, pistillate sepals with apical glands and ciliate with glandular trichomes ( Fig. 2 I View FIGURE 2 ) — an exclusive character combination among South American Tragia species.
Distribution and ecology:— Tragia peltata occurs in lowland evergreen forests in the Atlantic Forest biome ( Fig. 4 A View FIGURE 4 ), in forest interiors, and along the edges of forests adjacent to areas that have suffered recent disturbances. Until recently, the species was considered endemic to Ilha Grande Bay in Rio de Janeiro State ( Secco et al. 2020), but new populations were found during this study in the northern region of Espírito Santo State and in the southern region of Bahia State, expanding its area of occurrence ( Fig. 4 A View FIGURE 4 ).
The different regions where T. peltata occurs have different biogeographies. Fiaschi & Pirani (2009) divided the Atlantic Forest into two parts: a northern block (extending from latitude -5° in Rio Grande do Norte State to latitude -19° in northern Espírito Santo) in areas that share similarities with Amazonian vegetation; the southern block occurs from latitude -19° to latitude -29° (in Santa Catarina State) and shares similarities with the Andean subtropical flora.
As the two Atlantic Forest blocks are roughly divided by the Vale do Rio Doce region ( Fiaschi & Pirani 2009), T. peltata populations likewise appear to be separated by that geographic barrier. The role of the Rio Doce as a barrier between populations has been reported in the literature, and is sometimes considered a speciation mechanism ( Maciel et al. 2019, Santos et al. 2018). Additionally, although the known occurrences of T. peltata are in dense ombrophilous forest vegetation ( Fig. 4 B View FIGURE 4 ), they are separated by the semideciduous seasonal forest that occurs from southern Espírito Santo to northern Rio de Janeiro ( Fig. 4 B View FIGURE 4 ), which appears as another barrier between populations.
Phenology:— Tragia peltata was encountered flowering and fruiting in April and May and from September to December. Flower maturation apparently varies according to sex. The pistillate flowers of the inflorescence open while the staminate buds are still developing, and the staminate flowers open when fruits begin to develop, which sometimes creates an overlap. Due to this reproductive strategy, Vellozo (1881) and Müller Argoviensis (1874) had only fruiting material to describe T. peltata , making this the first description of staminate and pistillate flowers.
Conservation status:— Tragia peltata has an Area of Occupancy of 16,000 km 2 and an Extent of Occurrence 46,562.074 km ² which classifies it as Least Concern according to IUCN (2019). However, the conditions a(iii) and b (iii) of the B criteria may be applied. Most collections (10) of the T. peltata that was found on the Reserva Biológica de Sooretama (RBS) and Reserva Natural da Vale (RNV), which are two parts of a continuous forest among the municipalities of the Linhares and Sooretama on Espírito Santo state. Two other collections of T. peltata were made: a) on municipality of Itamaraju-BA, ca. 220 km from the forest of the RBS and RNV; b) on municipality of Angra dos Reis-RJ, ca. 620 km from the forest of the RBS and RNV ( Fig. 4 A View FIGURE 4 ).
The populations of T. peltata occur in areas of Atlantic Forest that have suffered historical fragmentation processes. Currently, the Atlantic Forest only have 11.26% of the original cover ( SOS Mata Atlântica 2008), and 80% of the fragments occupy less than 50 ha. The maintenance of those fragments is important for reducing isolation between populations. However, recent studies have documented their continuing size reductions ( Ribeiro et al. 2009) – which can expose T. peltata to risk of extinction.
Typification:— The typification of Vellozo’s names based on exsiccates is often impossible due to the lack of information concerning the locations of type collections ( Kury 2015, Milward-de-Azevedo 2017). Therefore, in accordance with articles 9.3, 9.4 and 9.11 of the International Code of Nomenclature for Algae, Fungi and Plants ( Turland et al. 2018), the original illustration n. 6 ( Vellozo 1831b) ( Fig. 1 View FIGURE 1 ) preserved in the Manuscript section of the Library of the National Museum of Rio de Janeiro [cat. no: mss1198659_010], and published in Vellozo, Fl. Flumin. 10, t. 6, 1831, iss selected as the lectotype.
Cardinal-McTeague, W. M. & Gillespie, L. J. (2016) Molecular phylogeny and pollen evolution of Euphorbiaceae tribe Plukenetieae. Systematic Botany 41: 329 - 347. https: // doi. org / 10.1600 / 036364416 X 691759
Cordeiro, W. P. F. S., Athie-Souza, S. M., Melo, A. L. & Sales, M. F. (2020 a) A new endangered species of Tragia (Euphorbiaceae) from the Brazilian Atlantic Forest. Systematic Botany 45: 839 - 844. https: // doi. org / 10.1600 / 036364420 X 16033962925358
Cordeiro, W. P. F. S., Athie-Souza, S. M., Torres, A. L., Melo, A. L. & Sales, M. F. (2020 b) A new record for Guyana reveals the disjunct distribution fo Tragia cearensis Pax & K. Hoffm. (Euphorbiaceae). Check List 16: 1603 - 1608. https: // doi. org / 10.15560 / 16.6.1603
Fiaschi, P. & Pirani, J. R. (2009) Review of plant biogeographic studies in Brazil. Journal of Systematics and Evolution 47: 477 - 496. https: // doi. org / 10.1111 / j. 1759 - 6831.2009.00046. x
Gutierrez de Sanguinetti, M. M. & Mulgura de Romero., M. E. (1986) Uma nueva especie de Tragia (Euphorbiaceae). Darwiniana 27: 491 - 497.
IUCN (2019) Guidelines for Using the IUCN Red List Categories and Criteria, version 14. Prepared by the Standards and Petitions Committee. Downloadable from: https: // nc. iucnredlist. org / redlist / content / attachment _ files / RedListGuidelines. pdf (accessed May 2020).
Kury, L. B. (2015) O naturalista Veloso. Revista de Historia 172: 243 - 277. https: // doi. org / 10.11606 / issn. 2316 - 9141. rh. 2015.98752
Linnaeus, C. (1753) Species Plantarum. Laurentius Salvius, Stockholm, 1200 pp.
Maciel, J. R., Souza, G. M., Wanderley, M. G. L., Zizka, G. & Alves, M. (2019) A new genus of Bromeliaceae endemic to Brazilian Atlantic Forest. Systematic Botany 44: 519 - 535. https: // doi. org / 10.1600 / 036364419 X 15620113920572
Milward-de-Azevedo, M. A. (2017) Typifications in Passiflora L. (Passifloraceae) described by Frei Jose Mariano da Conceicao Vellozo. Acta Botanica Brasilica 31: 613 - 618. https: // doi. org / 10.1590 / 0102 - 33062017 abb 0128
Muller Argoviensis, J. (1866) Euphorbiaceae excl. Euphorbieae. In: Candolle, A. L. P. P. de (ed.) Prodromus Systematis naturalis Regni Vegetabilis 15 (2). Masson, Paris, pp. 190 - 1286.
Muller Argoviensis, J. (1874) Euphorbiaceae .. In: Martius, C. F. P. & Eichler, A. W. (eds.) Flora Brasiliensis 11 (2). F. Fleischer, Leipzig, Germany, 752 pp., 104 pl.
Pax, F. & Hoffmann, K. (1919) Euphorbiaceae - Acalypheae - Plukenetiinae. In: Engler, A. (ed.) Das Pflanzenreich IV. 147. XI (Heft 68). W. Engelmann, Leipzig, pp. 1 - 108.
Ribeiro, M. C., Metzger, J. P., Martencen, A. C., Ponzoni, F. J. & Hirota, M. K. (2009) The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation 142: 1141 - 1153. https: // doi. org / 10.1016 / j. biocon. 2009.02.021
Rusby, H. H. (1907) An enumeration of the plants collected in Bolivia by Miguel Bang, Part 4. Bulletin of the New York Botanical Garden 4: 309 - 470.
Santos, B. F., Scherrer, M. V. & Loss, A. C. (2018) Neither barriers nor refugia explain genetic structure in a major biogeographic break: phylogeography of praying mantises in the Brazilian Atlantic Forest. Mitochondrial DNA Part A 29 (8): 1284 - 1292. https: // doi. org / 10.1080 / 24701394.2018.1445242
Secco, R. S., Cordeiro, W. P. F. S., Bigio, N. C., Pereira-Silva, R. A., Melo, A. L., Gama, B. R. A., Athie-Souza, S. M., Oliveira, J. C. P., Mendes, J. C. R., Leal, B. A., Silva, D. F. & Sales, M. F. (2020) Tragia. In: Brazil Flora Group 2021. Jardim Botanico do Rio de Janeiro. Available from: http: // floradobrasil. jbrj. gov. br / reflora / floradobrasil / FB 17691 (accessed 27 April 2021).
SOS Mata Atlantica (2008) Atlas dos remanescentes florestais da Mata Atlantica, periodo de 2000 a 2005. Fundacao SOS Mata Atlantica, Sao Paulo & Instituto Nacional de Pesquisas Espaciais INPE, Sao Jose dos Campos. Available at: http: // www. sosmatatlantica. org. br (accessed 27 April 2021).
Turland, N. J., Wiersema, J. H., Barrie, F. R., Greuter, W., Hawksworth, D. L., Herendeen, P. S., Knapp, S., Kusber, W. - H., Li, D. - Z., Marhold, K., May, T. W., McNeill, J., Monro, A. M., Prado, J., Price, M. J. & Smith, G. F. (2018) International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code). Regnum Vegetabile 159. Glashutten: Koeltz Botanical Books. https: // doi. org / 10.12705 / Code. 2018
Vellozo, J. M. C. (1831 b [1827]) Flora Fluminensis Icones 10. Lithogr. Senefelder, Paris, 119 pp.
Vellozo, J. M. C. (1881) Flora Fluminensis. Archivos do Museu Nacional do Rio de Janeiro, 461 pp.

FIGURE 1. Lectotype of Tragia peltata. Original plate of T. peltata (1831b) on Flora Fluminensis in the Manuscripts Section of the National Library of Rio de Janeiro (cat. no.: mss1198659_010).
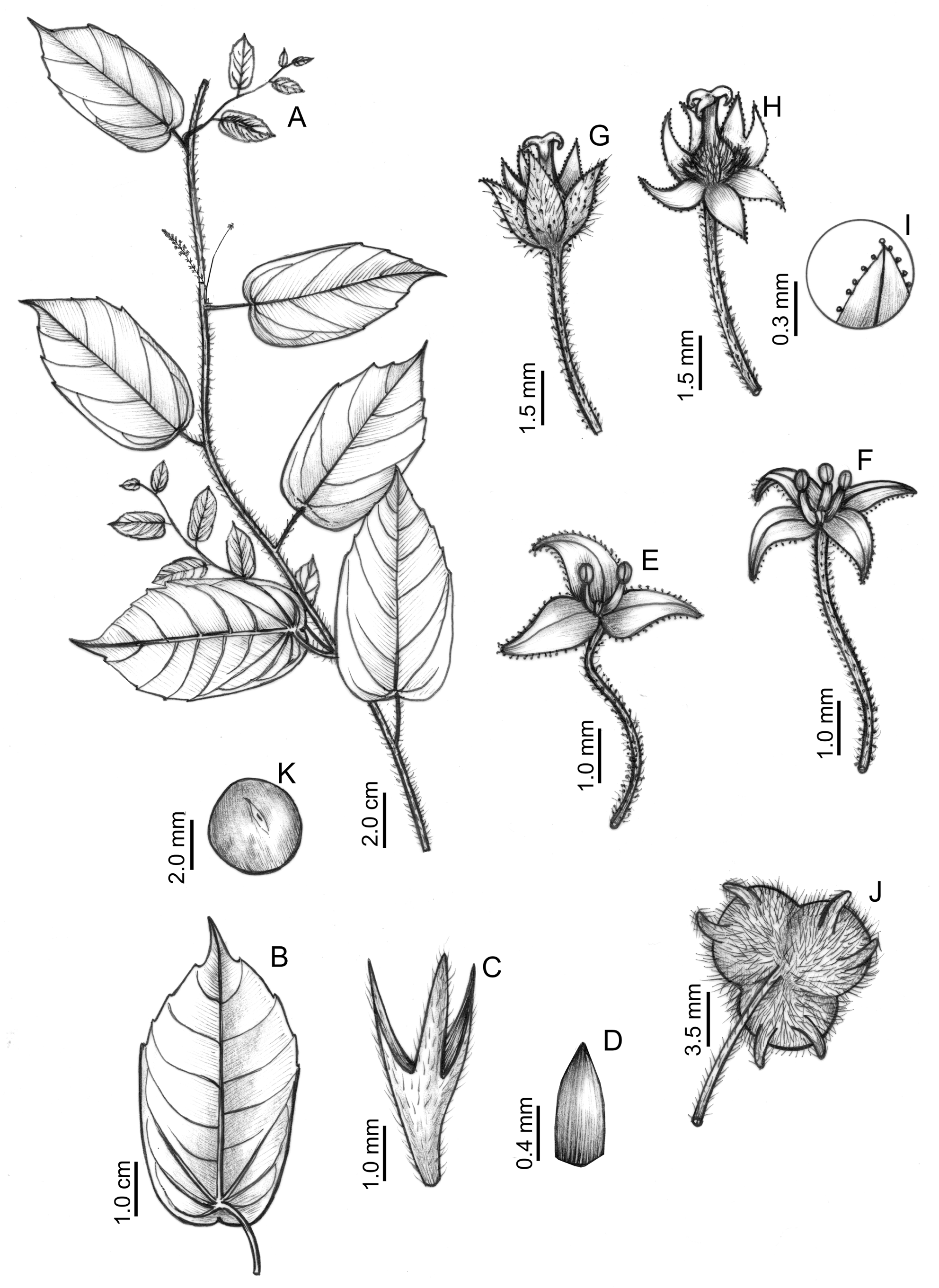
FIGURE 2. Tragia peltata. A. Flowering branch, B. Abaxial leaf face, C. Basal bract of the inflorescence. D. Staminate bract, E‒F. Staminate flowers, G–H. Pistillate flowers, I. Pistillate sepal, detail of the apex and ciliate margins with stipitate glandular trichomes, J. Fruit, K. Seed. Illustrated by Regina Carvalho, from H.F. Talbot s.n. [K 001205063] (A, B), D.A. Folli s.n. [IPA 66670] (C‒K).

FIGURE 3. Leaf variability in Tragia peltata. Population from Rio de Janeiro state. A. Leaf blade, B. Leaf base on adaxial view, C. Leaf margin. Population from Bahia and Espírito Santo states. D. Leaf blade, E. Base of leaf blade with petiole insertion close to the margin, F. Leaf margin. A‒C from D. Sucre & J. Gomes 11277 (RB); D‒F from T.S. Santos 1551 (CEPEC). Scale = 1 cm (A‒B and D‒E) or 0.1 cm (C and F).
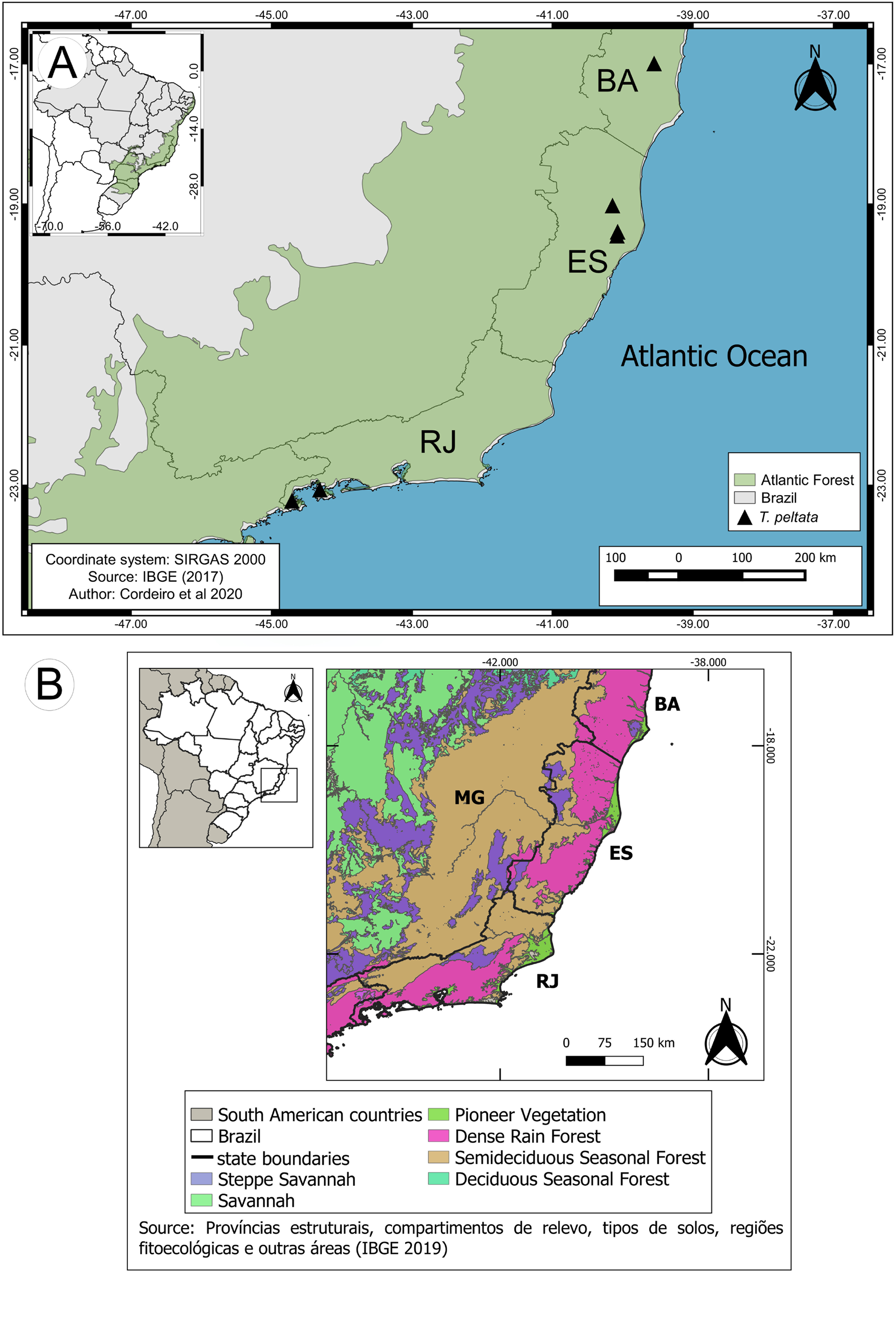
FIGURE 4. A. Geographic distribution of Tragia peltata in the Atlantic Forest in the states of Bahia (BA), Espírito Santo (ES), and Rio de Janeiro (RJ), Brazil. B. Spatial distribution of the Phytophisiognomies in the area of occurrence of T. peltata, stats of Bahia (BA), Espírito Santo (ES), Minas Gerais (MG), and Rio de Janeiro (RJ).
TABLE 1. Morphological differences among T. peltata and species with long pistillate pedicels.
| T. aurea | T. giardelliae | T. hoffmanniae | T. peltata | T. volubilis | ||
|---|---|---|---|---|---|---|
| General | Habit | Vine up to 2 m long | Vine up to 90 cm long | Vine, unknown length | Vine, unknown length | Subshrub volubilis up to 3 m long |
| aspect | Indument color in dry plants | Rust-coloured | White | White | White | White |
| Size | 1.5–3 × 0.4–0.5 mm | 4–4.5 × 2–2.5 mm | 3 × 0.5 mm | 2–3(–4) × 1–2 mm | 1–4 × 1–2 mm | |
| Stipules | Shape | Narrowly triangular | Ovate | Narrowly triangular | Triangular | Triangular-ovate |
| Persistence | Persistent | Deciduous | Deciduous | Deciduous | Persistent | |
| Petiole (length) | 5 mm | (3–)4–11 cm | 1–2 cm | 2–4 cm | 1–5 cm | |
| Leaf blade size | 2.5–5 × 0.7–2 cm | (4–)7–15 × (3.5–)5– 11 cm | 8–14 × 3.5–6 cm | 8.5–14.5 × 4–7 cm | 2–9 × 1–5.5 cm | |
| Leaf blade shape | Ovate | Broadly ovate | Elliptical or lanceolate | Ovate to oblong | Triangular-ovate, lanceolate or oblong | |
| Leaves | Base shape | Slightly cordate | Deeply cordate | Cordate or rounded | Emarginate to retuse-cordate, peltate | Cordate, obtuse, truncate or rounded |
| Apex shape | Acute or acuminate | Acuminate | Caudate | Acuminate | Acute or acuminate | |
| Leaf margin | Serrate | Dentate-acuminate | Entire | Sparsely serrate or subentire | Serrate or subentire | |
| Number of sepals | – | 3 | 3 | 3 or 4 | 3 | |
| Staminate | Sepal length | – | 1.2–1.6 mm | 2.5–2 mm | 1–1.2 mm | 0.9–1.5 mm |
| flowers | Sepal shape | – | Subcircular | Ovate | Elliptical | Obovate or elliptical |
| Number of stamens | – | 2 | 3 | 2–3 | 2–3 | |
| Number of sepals | 5 | 6 | 5 | 6(–8) | 6 | |
| Pistillate flowers | Number of sepal series Sepal length Sepal shape | 1 1.5 mm Lanceolate, apex obtuse | 2 3–3.5 mm Broadly elliptical, apex acute | 1 2.5 mm Narrowly triangular, apex acute | 1 2.5 mm Ovate, apex acuminate | 1 1–2.5 mm Ovate or lanceolate, apex acute |
| Apical gland | ? | Absent | Absent | Present | Absent |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |