
Laubierpholoe massiliana Zhadan, 2023
|
publication ID |
https://doi.org/10.5852/ejt.2023.875.2139 |
|
publication LSID |
lsid:zoobank.org:pub:6203D3DD-4A9B-4FFB-905D-ECAFB987792A |
|
persistent identifier |
https://treatment.plazi.org/id/446EA92C-FFC9-FF85-055C-CF3DFE2CFBEE |
|
treatment provided by |
Felipe (2023-06-16 13:02:59, last updated 2023-06-16 13:08:47) |
|
scientific name |
Laubierpholoe massiliana Zhadan |
| status |
sp. nov. |
Laubierpholoe massiliana Zhadan sp. nov.
urn:lsid:zoobank.org:act:4D95E54D-F91E-409C-AA71-74563AD97E9A
Figs 1–6 View Fig View Fig View Fig View Fig View Fig View Fig
Diagnosis
Body with 16–19 segments, 8–10 pairs of elytra; two pairs of closely arranged eyes; dorsal tentacular cirri of same length or slightly longer than ventral ones; notopodia smaller and shorter than neuropodia; notochaetae few (3–6), some neurochaetae with bidentate tips.
Etymology
The species name refers to the type locality (Massilia – the old Roman name for Marseille).
Material examined
All material investigated was collected in the Mediterranean Sea, Gulf of Lion, The Calanques, near Marseille ( Fig. 1B View Fig –С). The coordinates of the entrance of the Jarre Cave: 43.19556° N, 5.3658333° E; for 3PP Cave: 43.16306° N, 5.6° E.
Holotype
FRANCE • 1 specimen (pharynx everted, anal cirri damaged; 17 segments; body length without pharynx and anal cirri 0.925 mm, including pharynx 1.31 mm, body width 150 µm, including elytra 490 µm, including chaetae 720 µm); Mediterranean Sea , Gulf of Lion , The Calanques , near Marseille, Jarre Cave, deep part; 43.19556° N; 5.3658333° E; depth 18 m; 24 Oct 2019; P. Chevaldonné leg.; SCUBA-diving; sampling box; silty sand; mesh size 130 µm; preservation in glutaraldehyde 2.5% in PBS, storage in ethanol 70%; field number Ma 19-28-03; ZMMSU WS16462 . GoogleMaps
Paratypes
FRANCE • 15 specimens; same data as for holotype; preservation glutaraldehyde 2.5% in PBS, storage in ethanol 70% (7 specimens), SEM stubs (3 specimens), permanent slides (5 specimens); field number Ma 19-28-03; ZMMSU WS14001 GoogleMaps • 6 specimens; same locality as for holotype; 12 Mar. 2020; P. Chevaldonné leg.; SCUBA-diving; sampling box; mesh size 130 µm; field number Ma 20-03-01; preservation in glutaraldehyde 2.5% in PBS, SEM stubs; ZMMSU WS16511 GoogleMaps • 3 specimens; Jarre Cave , middle part; 10 May 2019; depth 19 m; P. Chevaldonné leg.; SCUBA-diving; sampling box; sandy silt; mesh size 130 µm; preservation in EtOH 96%; field number Ma 19-24-01; ZMMSU WS12418 • 3 specimens; 3PP Cave , middle part; 43.16306° N, 5.6° E; 28 Oct. 2019; depth 25 m; P. Chevaldonné leg.; SCUBA-diving; sampling box; clayey silt; mesh size 130 µm; preservation in glutaraldehyde 2.5% in PBS, storage in ethanol 70%; field number Ma 19-29-11; MNHN IA-TYPE 2058 GoogleMaps • 2 specimens; same data as for preceding; SEM stubs; ZMMSU WS13977 GoogleMaps .
Other material
FRANCE • 6 specimens; 3PP Cave , middle part; 43.16306° N, 5.6° E; 10 May 2019; depth 25 m; P. Chevaldonné leg.; SCUBA-diving; sampling box; clayey silt; EtOH 96% (4 specimens), SEM stubs (2 specimens); field number MA19-20-01; ZMMSU WS12292 GoogleMaps • 1 specimen; 3PP Cave , middle part; 43.16306° N, 5.6° E; depth 25 m; 7 May 2019; P. Chevaldonné leg.; SCUBA-diving; hand corer; clayey silt; mesh size 130 µm; DNA; field number Ma 19-13-01; ZMMSU WS12216 GoogleMaps ; • 1 specimen; same locality as for preceding; 10 May 2019; P. Chevaldonné leg.; SCUBA-diving; sampling box; clayey silt; mesh size 130 µm; DNA; field number Ma 19-20-01; ZMMSU WS12619 GoogleMaps • 1 specimen; 10 May 2019; 3PP Cave , deep part; 43.16306° N; 5.6° E; depth 25 m; P. Chevaldonné leg.; SCUBA-diving; sampling box; clayey silt; mesh size 130 µm; DNA; field number Ma 19-19-12; ZMMSU WS12617 GoogleMaps • 1 specimen; Jarre Cave , middle; 43.19556° N; 5.36583° E; depth 19 m; 10 May 2019; P. Chevaldonné leg.; SCUBA-diving; sampling box; sandy silt; mesh size 130 µm; DNA; field number Ma 19-24-01; ZMMSU WS12437 GoogleMaps .
Description (based on all specimens investigated)
BODY. Short, up to 1.2 mm long (without appendages), up to 0.25 mm wide (without elytra), 0.65 mm (with elytra), 0.8 mm (with chaetae), 16–19 segments, body surface smooth. Living worms whitish, semi-transparent, often with transverse white stripe along anterior border of prostomium, intestine content yellowish to brown ( Fig. 2A–B View Fig ). Elytra 8–10 pairs, round, transparent, with few oval to cirriform papillae on surface and along lateral and posterior borders; some enclosing developing embryos ( Figs 2A–C, E View Fig , 3A, C View Fig ). Prostomium and tentacular segment fused; prostomium oval to trapezoidal, bilobed; lobes rounded with notches above tentaculophores ( Figs 3D–G View Fig , 5A View Fig ). Two pairs of very closely situated, almost fused eyes near anterolateral border, anterior pair larger than posterior ( Figs 2A–F View Fig , 5A View Fig ). Median antenna with large ceratophore and short style, dorsal in anterior notch of prostomium ( Figs 3D–G View Fig , 5A View Fig ). Tentacular segment bearing medial anterior horns and lateral tentaculophores with dorsal and ventral tentacular cirri; tentacular cirri longer than median antenna, ventral tentacular cirrus equal or subequal to dorsal cirrus ( Figs 3D–G View Fig , 5A View Fig ). Anterior horns conical, arising medially from ventral tentacular cirri ( Figs 3D–G View Fig , 5A View Fig ). Upper and lower lips bearing conical papillae with a single sensory cilium distally ( Figs 3E–G View Fig , 4A View Fig ). Palps very long, up to 250 µm, stout, tapering, ventrolateral to tentaculophores ( Figs 2A, D–G View Fig , 3D–E, G View Fig , 5A View Fig ). Segment 2 with first pair of bulbous elytrophores, biramous parapodia, ventral buccal cirri lateral to mouth, longer than dorsal tentacular cirri ( Figs 3D–G View Fig , 5A View Fig ), ventral cirri from segment 3 onwards smaller than buccal cirri ( Fig. 4A View Fig ). Pharynx reaching segment 7 when inverted, with two pairs of jaws, 9 dorsal and 9 similar-sized ventral border papillae or three median papillae slightly smaller than lateral ( Figs 2C–D, F–H View Fig , 4B View Fig , 5D View Fig ).
PARAPODIA. Long, notopodia smaller and shorter than neuropodia, both with conical acicular lobes with projecting acicula; notopodia with two ciliated strips on dorsal side, one near base, the second subdistally ( Figs 4C–D View Fig , 5C View Fig ). Notochaetae few (3–6), slender, tapering to fine tips, straight or slightly curved, with series of small denticles ( Figs 4C–D View Fig , 5B–C View Fig ). Neurochaetae longer and stouter than notochaetae, compound ( Figs 2D–G View Fig , 3A–B View Fig ); shafts with long distal spines; upper blades slightly longer than lower; blades serrated, straight, falcate and of two types: supraacicular blades unidentate and subacicular blades uni- and bidentate (bidentate tips distinct at high magnification, Figs 4C–G View Fig , 5B View Fig ). Ventral cirri at neuropodial bases, thin, short, with round tips ( Figs 4A View Fig , 5C View Fig ). Pygidium with pair of very long anal cirri, up to 550 µm ( Fig. 2A, C View Fig ), easily lost; anus terminal.
Type locality
The Calanques, near Marseille, Jarre Cave ( Fig. 1C View Fig ).
Distribution
The Calanques, near Marseille, Jarre and 3PP marine caves.
Ecology
Inhabits the upper layer of soft sediments in the middle and deep parts of marine caves at a depth of 19– 25 m. The sediment type in Jarre Cave was defined as silty sand in the deep part and sandy silt in the middle part, and in 3PP Cave as clayey silt in both deep and middle parts ( Table 2).
Molecular data
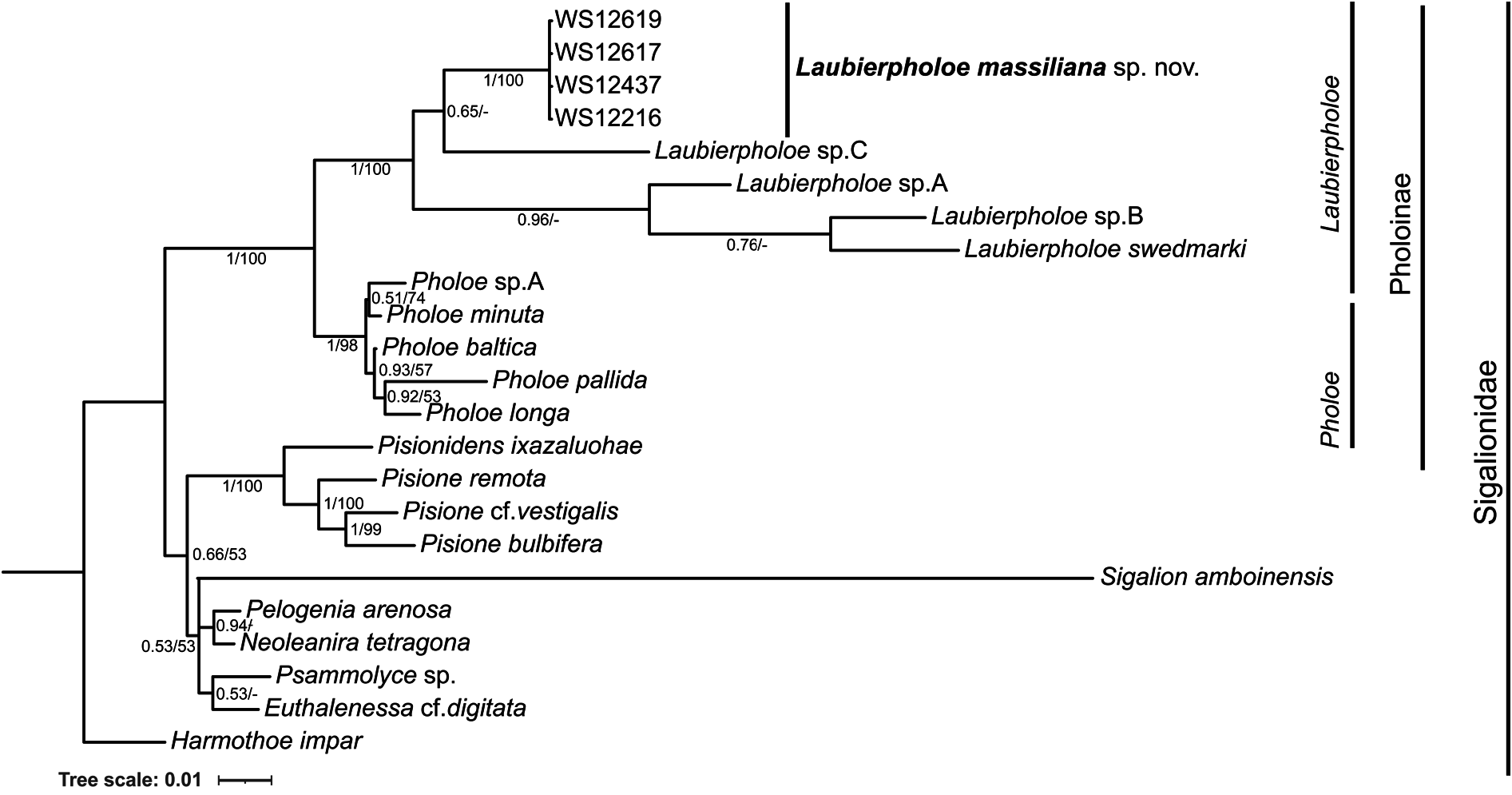
The trees obtained with the 18S rRNA and 28S rRNA concatenated dataset with BI ( Fig. 6 View Fig ) and ML analyses (Supp. file 1) have similar topology. Analysed specimens of Laubierpholoe massiliana sp. nov. form a highly supported clade within Laubierpholoe (PP = 1, B = 100); their sequences are identical despite their origin from two different caves. Laubierpholoe (PP = 1, B = 97), Pholoe (PP = 1, B = 98) and Pholoinae (PP = 1, B = 100) are well-supported groups within Sigalionidae . Laubierpholoe includes L. massiliana sp. nov. and all other species present in GenBank ( L. swedmarki and undescribed species A, B and C); it forms a sister group with Pholoe . The relationships of species of Laubierpholoe are different in BI and ML analyses and the subclades have low support. In the BI tree, L. massiliana sp. nov. is in a sister group with Laubierpholoe sp. C , and together they form a sister group with Laubierpholoe sp. A , Laubierpholoe sp. B and L. swedmarki ; Laubierpholoe sp. B and L. swedmarki form a clade which is a sister group with Laubierpholoe sp. A . In the ML tree, L. massiliana sp. nov. is in a sister group with the clade comprising Laubierpholoe sp. B and Laubierpholoe sp. C , and together they form a sister group with the clade L. swedmarki – Laubierpholoe sp. A .
Key to species of Laubierpholoe
1. Eyes absent; notopodium about as long as neuropodium ......................... L. maryae Pettibone, 1992 – Eyes present ...................................................................................................................................... 2
2. Dorsal tentacular cirri of same length or slightly longer than ventral; some neurochaetae with bidentate tips (look under high magnification) ................................................... L. massiliana Zhadan sp. nov. – Dorsal tentacular cirri much longer than ventral, all neurochaetae unidentate ................................ 3
3. All neurochaetal blades similar, short ............................................................................................... 4 – Neurochaetal blades of different size and shape, can include spinigers, or smooth and serrate, or hook-like types .................................................................................................................................. 5
4. Dorsal tentacular cirrus of about same length as median antenna, eyes separate, notochaetae numerous (20–30) .................................................................................................. L. antipoda ( Hartman, 1967) – Dorsal tentacular cirrus twice as long as median antenna, eyes closely arranged, notochaetae few (2–4) .................................................................................................... L. swedmarki ( Laubier, 1975)
5. Notopodium longer than neuropodium, notochaetae numerous; neurochaetae of two types: supraacicular with blades long, tapering to capillary tips; subacicular with blades short .................. ...................................................................................................................... L. riseri Pettibone, 1992
– Notopodium shorter and smaller than neuropodium, 4–8 notochaetae; neurochaetae starting from CH 4 have serrated and smooth blades, posteriormost five segments also with hook-like neurochaetal blades ............................................................................................ L. indooceanica Westheide, 2001 .
Hartman O. 1967. Polychaetous annelids collected by the USNS Eltanin and Staten Island cruises, chiefly from Antarctic Seas. Allan Hancock Monographs in Marine Biology 2: 1 - 387.
Laubier L. 1975. Adaptations morphologiques et biologiques ches un aphroditien interstitiel: Pholoe swedmarki sp. n. Cahiers de Biologie marine 16 (311): 671 - 683.
Pettibone M. H. 1992. Contribution to the polychaete family Pholoidae Kinberg. Smithsonian Contributions to Zoology 532: 1 - 24. https: // doi. org / 10.5479 / si. 00810282.532
Westheide W. 2001. Laubierpholoe indooceanica, a new interstitial polychaete (Pholoidae) from South India and the Seychelles. Cahiers de Biologie marine 42 (4): 327 - 332. https: // doi. org / 10.21411 / CBM. A. F 7 F 42276
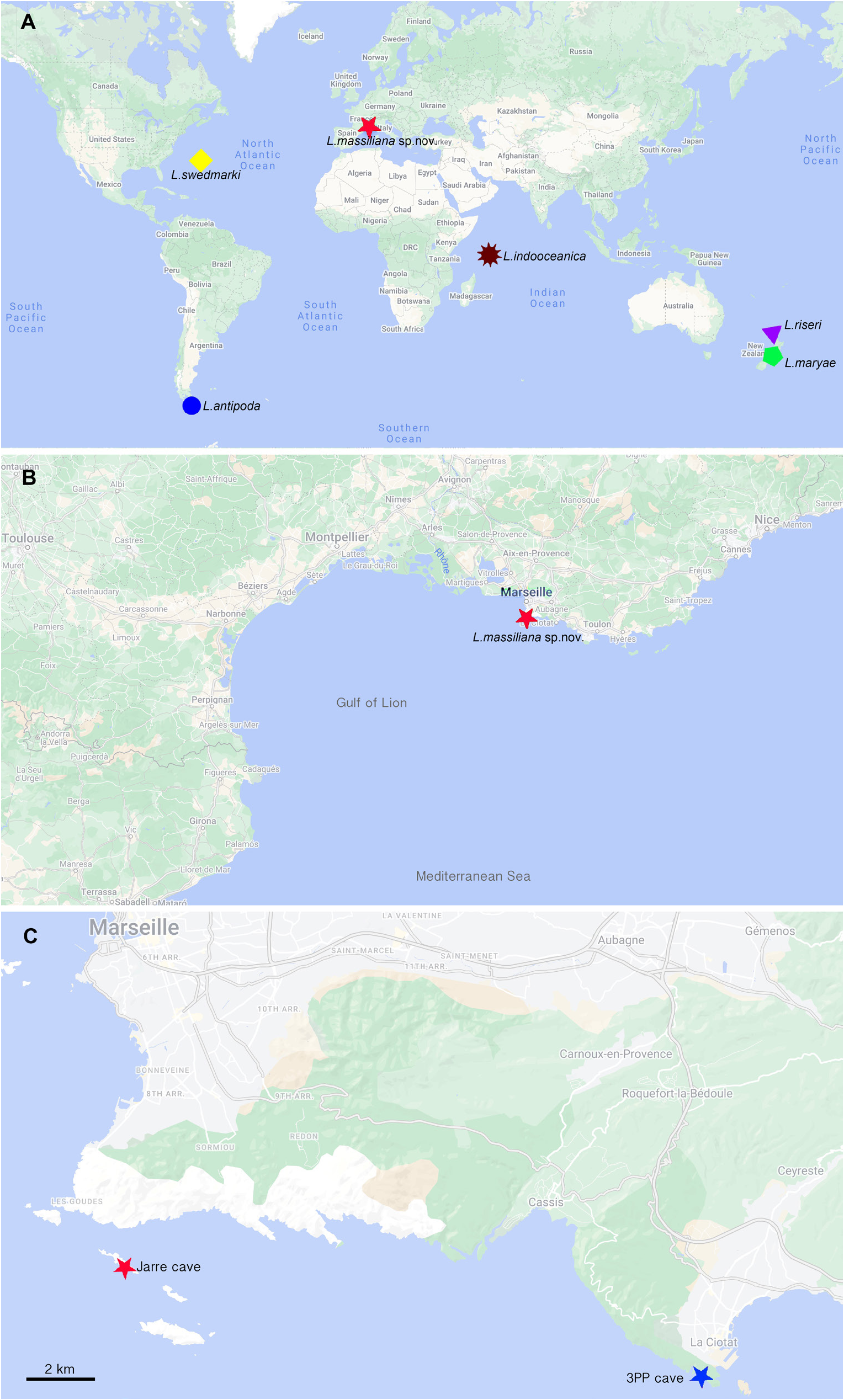
Fig. 1. A. World map showing type locations of all species of Laubierpholoe Pettibone, 1992. B. Type location of Laubierpholoe massiliana Zhadan sp. nov. C. Locations of the two caves where Laubierpholoe massiliana sp. nov. was sampled.

Fig. 2. Laubierpholoe massiliana Zhadan sp. nov., light microscopy. A–B. Living specimens from different samples. C. ZMMSU WS12418, paratype, general view with proboscis everted. D. ZMMSU WS16462, holotype, general view, proboscis everted. E. ZMMSU WS14001, paratype, general view. F–H. ZMMSU WS14001, paratype, compound microscope. F. General view. G. Anterior part, dark field. H. Right jaws.
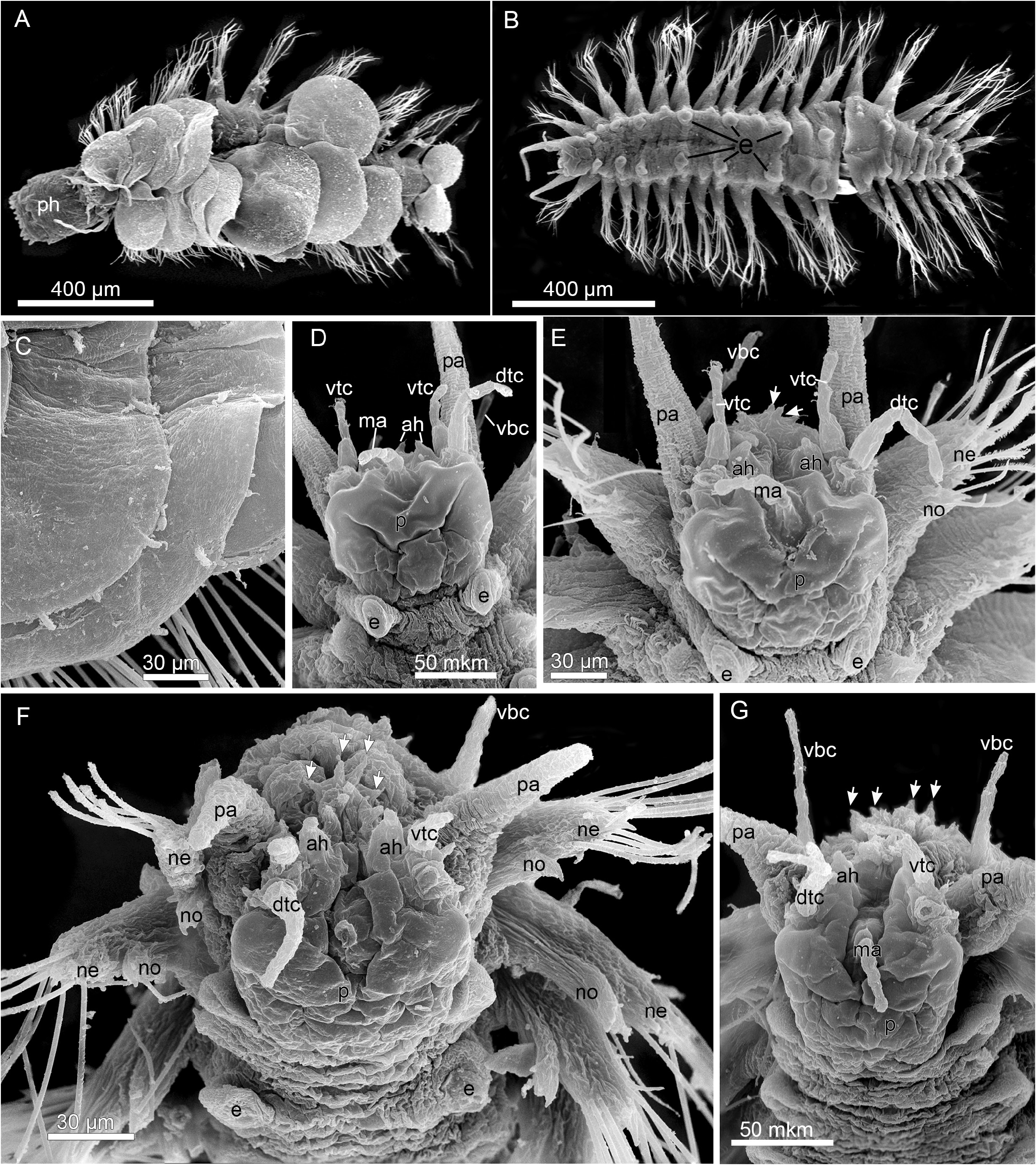
Fig. 3. Laubierpholoe massiliana Zhadan sp. nov., SEM. A. ZMMSU WS12292, general view.B. ZMMSU WS14001, general view, elytra omitted. C. ZMMSU WS13977, elytra. D. ZMMSU WS14001, anterior end, dorsal view. E. Same, dorso-anterior view. F. ZMMSU WS16511, anterior end, dorso-anterior view, median antenna broken. G. ZMMSU WS16511, dorso-anterior view. Abbreviations: ah = anterior horns; dtc = dorsal tentacular cirrus; e = elytrophores; ma = median antenna; ne = neuropodium; no = notopodium; p = prostomium; pa = palp; ph = pharynx; vbc = ventral buccal cirrus; vtc = ventral tentacular cirrus. Arrows indicate papillae.
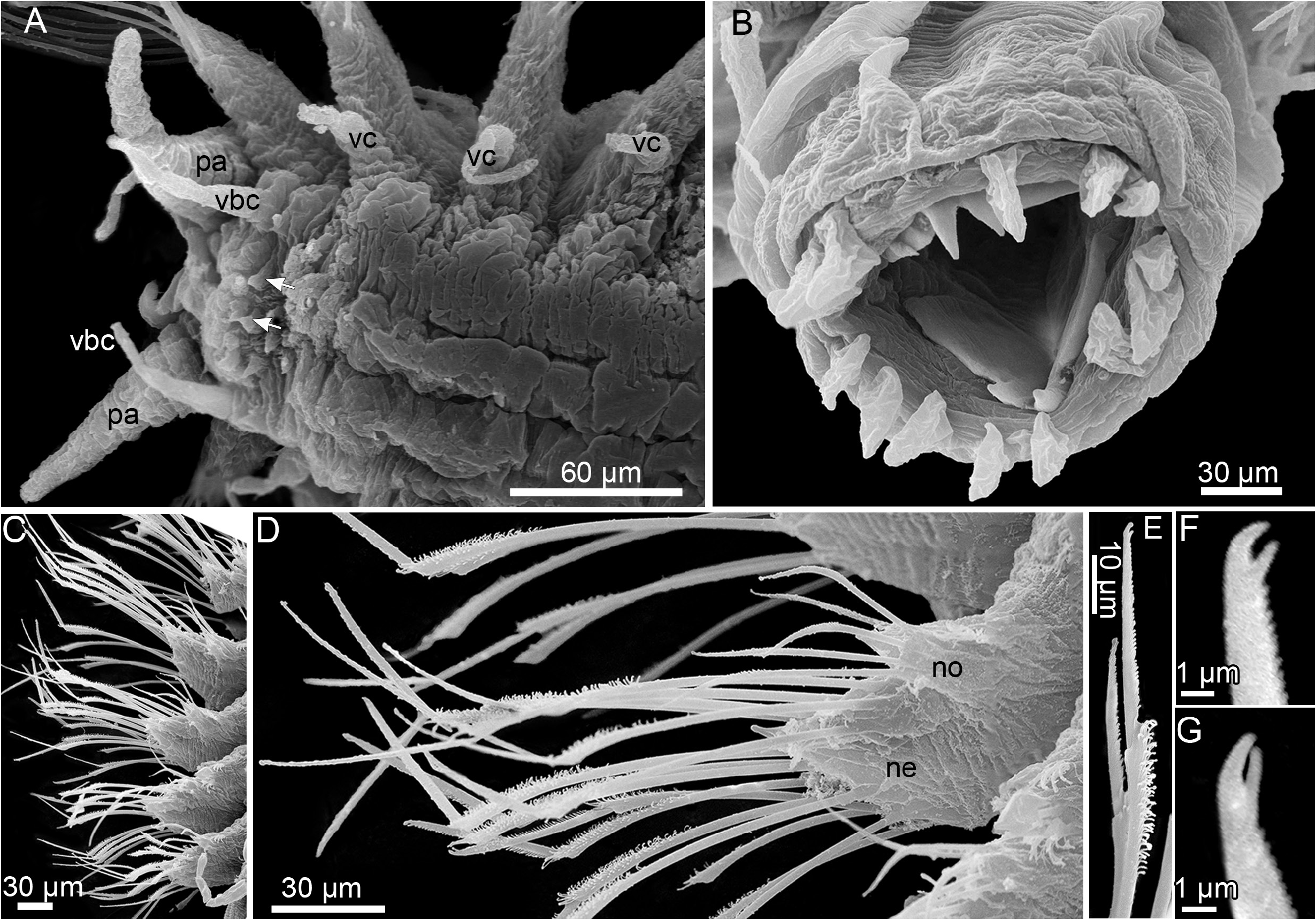
Fig. 4. Laubierpholoe massilianaZhadan sp. nov., SEM.A. ZMMSU WS16511, ventral view.B. ZMMSU WS16511, pharynx, dorso-anterior view.C. ZMMSU WS14001, parapodia of segments I-V, dorso-anterior view. D. Same, parapodia of segment III, anterior view. E. ZMMSU WS13977, bidentate neurochaetae. F–G. ZMMSU WS12292, tips of bidentate neurochaetae. Abbreviations: ne = neuropodium; no = notopodium; pa = palp; vbc = ventral buccal cirrus; vc = ventral cirrus. Arrows indicate papillae.

Fig. 5. Laubierpholoe massiliana Zhadan sp. nov., line drawings. A. Anterior end, dorso-anterior view. B. Notochaeta and bidentate neurochaeta. C. Parapodium, anterior view. D. Jaw. Abbreviations: ah = anterior horns; dtc = dorsal tentacular cirrus; ma = median antenna; ne = neuropodium; no = notopodium; p = prostomium; pa = palp; vbc = ventral buccal cirrus; vc = ventral cirrus; vtc = ventral tentacular cirrus. Arrows indicate papillae.
| PBS |
Chambers Institute, Tweeddale Museum |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
1 (by felipe, 2023-06-16 13:02:59)
2 (by ExternalLinkService, 2023-06-16 13:08:47)
3 (by juliana, 2023-06-19 13:26:37)
4 (by ExternalLinkService, 2023-06-19 13:36:44)
5 (by ExternalLinkService, 2023-06-19 15:01:52)
6 (by plazi, 2023-11-09 10:22:43)
7 (by ExternalLinkService, 2023-11-09 12:20:18)