
Philander quica ( Temminck, 1824 )
|
publication ID |
https://doi.org/ 10.1206/3891.1 |
|
persistent identifier |
https://treatment.plazi.org/id/36452319-FFA5-FFC9-11F5-FBC9FBDCFD42 |
|
treatment provided by |
Carolina |
|
scientific name |
Philander quica ( Temminck, 1824 ) |
| status |
|
Philander quica ( Temminck, 1824)
Didelphis quica Temminck, 1824: 36 ; type locality (fixed by lectotype selection; Hershkovitz, 1959: 342) “Sapitibi” (= Sepetiba at 22°58′ S, 43°42′ W; Paynter and Traylor, 1991), Rio de Janeiro, Brazil.
Metachirus opossum azaricus Thomas, 1923: 604 ; type locality “Sapucay” (= Sapucaí at 25°41′ S, 56°57′ W; Paynter, 1989), Paraguarí, Paraguay GoogleMaps .
TYPE MATERIAL: Temminck (1824: 36–38) based his description of Didelphis quica on an unknown number of specimens from various museums, presumably including one or more examples collected by Johann Natterer, whose information about the species was prominently acknowledged (“Nous devons à M. Natterer la connaissance plus exacte de cette espèce qu’il a envoyée au musée impérial de Vienne...”). In a published catalog of Natterer’s mammals, Pelzeln (1883: 110–111) listed two specimens of D. quica , male and female, collected in 1818 at “Sapitiba” (= Sepetiba) near Rio de Janeiro. Of these, Hershkovitz (1959) designated the female as lectotype, apparently sight unseen. Unfortunately, this specimen is no longer in Vienna, where only the male topotype ( NMW 7687/ST 1012) can now be found. The female may have been exchanged or gifted to Temminck, whose cabinet was subsequently transferred to the Leiden museum (formerly the Rijksmuseum van Natuurlijke Historie, now the Naturalis Biodiversity Center; S. Engelberger, personal commun., 26 November 2014), but no specimen currently in Leiden can be positively identified as Hershkovitz’s lectotype (S. van der Mije, 25 November 2014).
In the absence of any compelling evidence to the contrary, we accept Hershkovitz’s (1959) lectotype designation as valid. Although the specimen in question appears to have been lost (or to be unidentifiable), the fixation of the type locality is sufficient for confident application of Temminck’s epithet to the Atlantic Forest species of southeastern Brazil. The male topotype mentioned above (consisting of the mounted skin and extracted skull of a very old animal with much-faded pelage and teeth worn away almost to the roots) is not taxonomically informative, but other examined specimens of Philander from the Brazilian state of Rio de Janeiro (in the AMNH and ZMB; see below) exhibit all the diagnostic traits that we attribute to P.quica in the description that follows.
DISTRIBUTION AND SYMPATRY: Sequenced material and examined specimens that we assign to Philander quica are from rainforested tropical and subtropical landscapes in southeastern Brazil, northeastern Argentina (Misiones), and eastern Paraguay ( fig. 9 View FIG ). Although P. quica is the only species of Philander known to occur throughout this biome (the Mata Atlântica of Brazilian authors), it might occur sympatrically with P.canus along its margins, where Atlantic rainforests come into contact with (or grade into) the gallery formations and semideciduous forests apparently preferred by the latter species. Additionally, the range of P.quica might contact that of P. opossum somewhere along the coastline between Bahia and Rio Grande do Norte. 8 Published records of this species from the Cerrado, the Chaco, and western Amazonia (in Patton and da Silva, 1997, 2008; Hershkovitz, 1997) are based on misidentifications (see Remarks, below).
DESCRIPTION: Dorsal pelage short (usually <14 mm) and uniformly grayish, sometimes indistinctly darker along the midline but never with a distinctly blackish middorsal stripe ( fig 10 View FIG ); fur of crown (between the ears) usually grizzled gray but sometimes blackish; pale preau-
8 We are told (D. Astúa, personal commun.) that this is unlikely. The southeasternmost record we have seen of Philander opossum is a specimen from Canudos, Rio Grande do Norte, Brazil, collected by F. Lima in 1920 ( FMNH 24790 About FMNH ) .
, Minas Gerais (33): JQ778966 View Materials Brazil, Minas Gerais, Cruzeiro (30): LC4 , Rio de Janeiro (41): JQ7789700, Minas Gerais (31): YL181, Rio de Janeiro (42): U34679 View Materials , São Paulo (48): MAM211, Minas Gerais (32): JF281029 View Materials , Rio de Janeiro (43): MAM183, Rio de Janeiro (44): MCL137 Espírito Santo (25): GU112936 View Materials Espírito Santo (24): GU112940 View Materials Espírito Santo (24): MAM189 Espírito Santo (26): GU112941 View Materials Espírito Santo (23): GU112942 View Materials quica São Paulo (46): MAM41 São Paulo (47): JQ778968 View Materials Paraná (39): YL225 São Paulo (46): KJ868146 View Materials , Misiones (3): JQ778959 View Materials , Misiones (4): JQ778961 View Materials Paraná (40): LPC876 Paraná (39): JQ778969 View Materials Bahia (22): YL107 Paulo (45): MAM64 (40): GU112939 View Materials , Misiones (3): JQ778964 View Materials , Misiones (3): JQ778963 View Materials , Paraguaí (77): JQ778960 View Materials
ricular spot absent or indistinct; ventral fur pale, usually self-whitish or -yellowish, sometimes with broad lateral zones of gray-based hairs on the throat and between the fore- and hind legs, but apparently always self-colored in the midline ( fig. 11 View FIG ); pinnae pale (unpigmented) basally, but abruptly blackish distally; dorsal pelage of hind feet often indistinctly darker laterally than medially, but never distinctly blackish or with blackish markings; scaly part of tail usually onethird to slightly less than one-half white distally. Skull ( fig. 12A, D View FIG ) substantially smaller than those of most congeneric species (except P. canus ). Nasal bones neither conspicuously elongated nor very short (about 47% of condylobasal length on average), not extending posteriorly to or between postorbital processes. Third upper premolar (P3) labial cingulum incomplete, apparently never extending to anterior base of crown; crown length of upper molar series 12.3 ± 0.4 mm (sexes combined; observed range 11.4–13.2 mm, N = 40); enameled lingual surfaces of upper molars smooth, not crenulated; pre- and postcingula consistently absent; lower molar posterior cingulids absent.
PHYLOGEOGRAPHY AND GEOGRAPHIC VARIATION: Our phylogenetic analysis of 28 cytochrome- b sequences of Philander quica spanning some 10 degrees of latitude (from the Brazilian state of Bahia in the north to the Argentinian province of Misiones in the south) provides scant evidence of phylogeographic structure ( fig. 13 View FIG ). This lack of genetic differentiation with distance is accompanied by an absence of conspicuous geographic variation in morphology. In effect, this appears to be a genetically and phenotypically homogeneous taxon.
COMPARISONS: Philander quica closely resembles P.canus , which is similar in size ( tables 7, 8); also has uniformly grayish dorsal pelage and self-whitish, -yellowish, or -buffy ventral fur ( figs. 10 View FIG , 11 View FIG ); and is not visually distinctive in any aspect of cranial appearance ( fig. 12 View FIG ). Chemisquy and Flores (2012) suggested that these taxa could be distinguished by the width of the postorbital constriction (least postorbital breadth in our terminology), but the samples we measured exhibit broad overlap in this dimension (e.g., 7.8–8.8 mm in P.quica males versus 7.4–8.4 mm in P.canus males). Philander quica and P.canus also have broadly overlapping distributions in the plane of the first two principal components that we computed from craniodental measurements of both taxa (not shown), a result consistent with our impression that these species are metrically very similar. Instead, qualitative morphological comparisons are more informative.
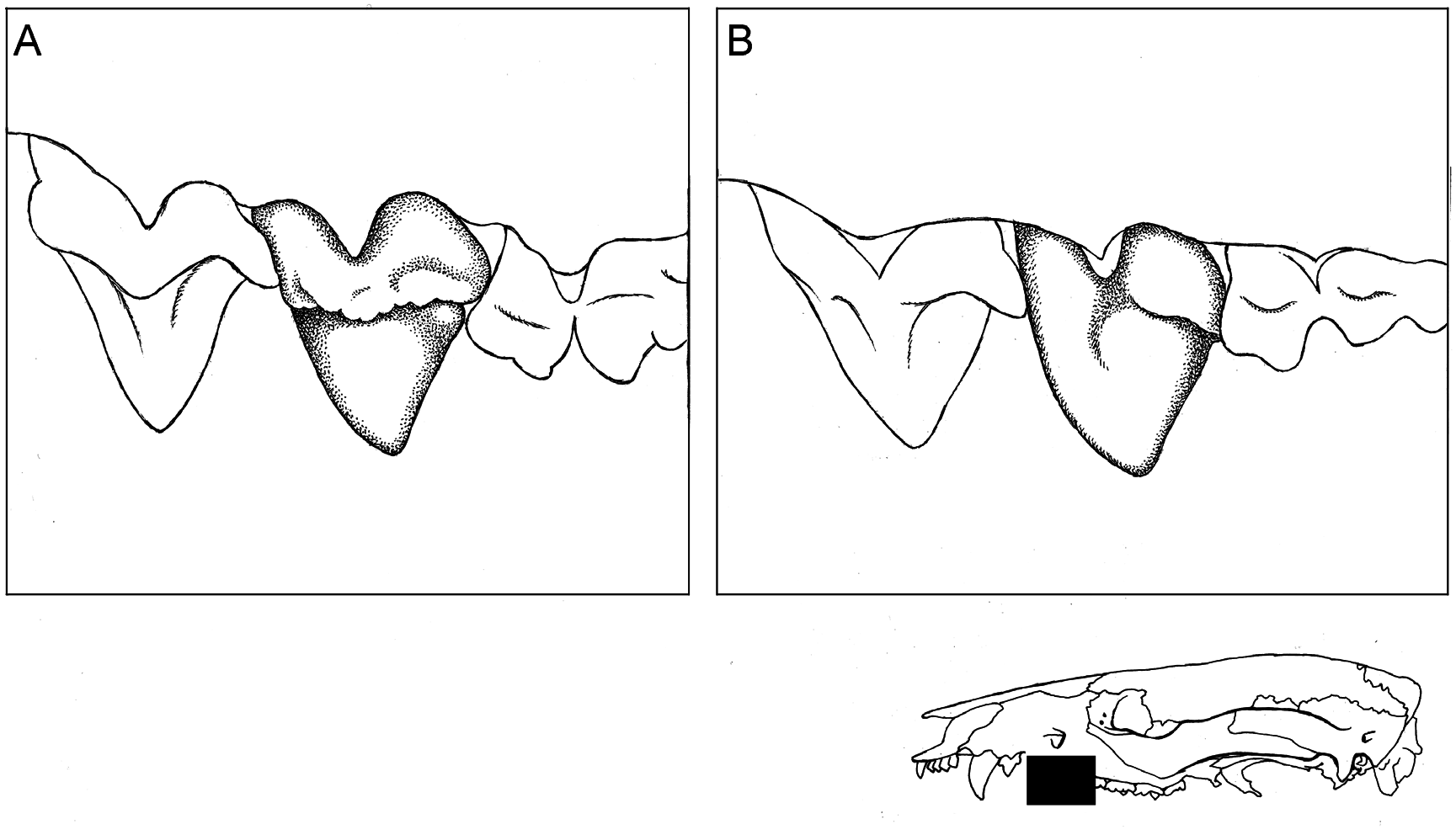
In side-by-side comparisons, the molars of Philander quica appear to have somewhat less well-developed anterolabial cingula, narrower protocones, deeper ectoflexi (especially on M3), and longer postmetacristae than those of P. canus , but the single most useful dental trait that distinguishes these taxa is the morphology of P3. Whereas the third upper premolar of P.canus always has a complete labial cingulum that extends along the entire base of the tooth from anterior to posterior ( fig. 14A View FIG ), the labial cingulum of P3 is incomplete in P.quica , apparently never extending anteriorly past the middle of that tooth ( fig. 14B View FIG ). Unfortunately, we have not found any external trait by which these species can be reliably identified in the field.
Close comparisons between Philander quica and our new species, P. pebas , seem unnecessary given their widely separated geographic distributions ( fig. 9 View FIG ), large genetic and morphometric distances (appendices 3, 4), and salient qualitative differences ( table 6).
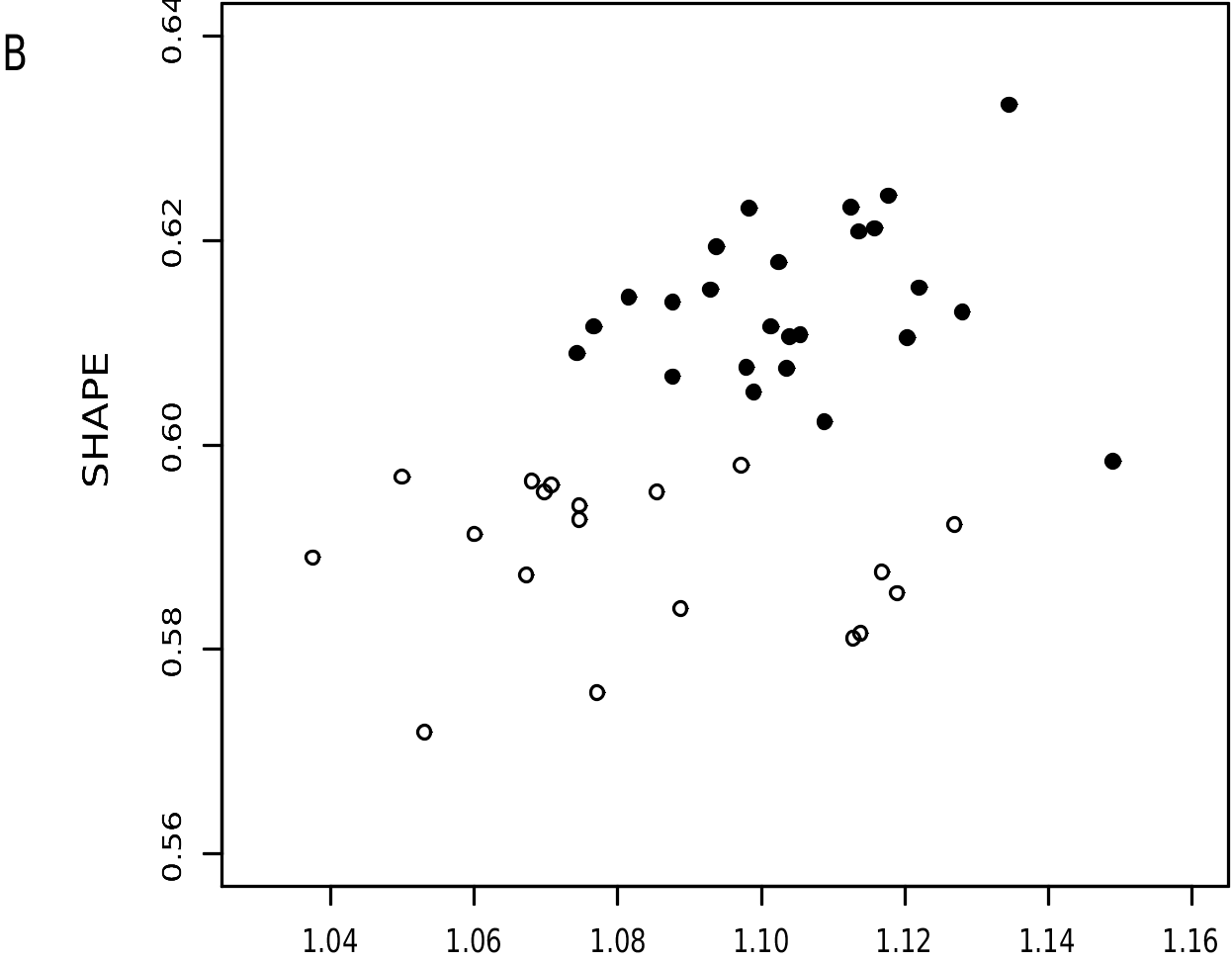
By contrast, Philander quica and P. opossum merit comparison because they are externally similar (with uniformly grayish dorsal fur and mostly self-colored ventral fur) and might eventually be found to occur sympatrically in eastern Brazil (see above). Although same-sex univariate comparisons ( table 9) reveal some overlap in all measured craniodental dimensions, principal-components analysis indicates that these species have discrete multivariate distributions ( fig. 15A View FIG ). Because the axis of species discrimination is approximately perpendicular to the more or less parallel axes of within-species variation, and because these axes are oblique to PC1 and PC2, we computed size and size-independent shape factors to obtain vectors with more interpretable coefficients ( fig. 15B View FIG ; table 10). The latter suggest that, independent of general-size allometries, P. opossum has longer but narrower nasals, a longer palate, and much larger molars than P. quica .
Differences in nasal shape between Philander quica and P. opossum are subtle but useful for visual identification of skulls: expressed as a percentage, the ratio NB/NL is about 25% on average in P. quica versus about 22% in P. opossum . Additionally, the shorter/broader nasals of P. quica never extend posteriorly to or between the postorbital processes, whereas the longer nasals of P. opossum often (in about two-thirds of examined specimens) extend to or between the postorbital processes. Whereas P. quica always has an incomplete labial cingulum on P3 ( fig. 14B View FIG ), the labial cingulum of P3 is narrowly complete (ending along the entire base of the tooth when unworn) in about 28% of examined specimens of P. opossum . Externally, P. opossum has better-defined preauricular spots and more saturated (consistently buffy) underparts than P. quica (which usually has whitish or yellowish ventral fur), and P. opossum tends to have a more extensively white-tipped tail (over half the specimens we examined have tails that are about ⅔ white) than P. quica (in which most specimens have tails that are ≤ ½ white). In side-by-side comparisons, the furred basal portion of the tail is visibly longer in P. opossum than in P. quica ; unfortunately, this trait is difficult to quantify due to the absence of a definite anterior landmark for relevant measurements.
REMARKS: Most recent authors have used the binomen Philander frenatus for this species following Patton and da Silva (1997), but the holotype of frenatus was collected in eastern Amazonia, and we treat that name as a junior synonym of P. opossum (see appendix 5).
PC1
SIZE
Hershkovitz (1997) used the trinomen Philander opossum quica for material that we refer to P. quica , P.canus , and P. pebas . He mapped the range of P. o. quica as including much of the Cerrado, Pantanal, and western Amazonia, but none of his material from Peru, Bolivia, or central Brazil corresponds to P.quica as recognized in this report. Hershkovitz (1997) regarded Didelphis myosuros Temminck, 1824 , as a synonym of P. o. quica and designated a lectotype for this purpose, but his lectotype designation is invalid because Pohle (1927) had previously designated a lectotype for D. myosuros , which is currently recognized (e.g., by Gardner and Dagosto, 2008) as a subspecies of Metachirus nudicaudatus (Geoffroy, 1803) .
According to Patton and da Silva (1997) this species—which they called Philander frenatus (see above)—occurs in the Brazilian state of Goiás, but they did not list any examined specimens from Goiás, and all the specimens of Philander that we examined from that state are unambiguously referable to P. canus (see below). Subsequently, Patton and da Silva (2008) mapped the range of P. “ frenatus ” as extending to the Chaco biome in the Argentinian province of Formosa, but the marginal record in question is based on AMNH 256980, a juvenile specimen that we reidentified as P. canus .
The status and relationships of Thomas’s azaricus has long been unsettled. Whereas Cabrera (1958) and Patton and da Silva (1997) treated this nominal taxon as a valid subspecies of P. opossum, Hershkovitz (1997) assigned it to the synonymy of P. o. quica , and Patton and da Silva (2008) included it in their synonymy for P. frenatus . Recently, Chemisquy and Flores (2012) analyzed a cytochrome- b sequence from a topotype of azaricus and found that it belonged to the Atlantic Forest haplotype group that they called P. frenatus . We examined the type series of azaricus and an additional 13 topotypes for this report; all of these specimens (21 in total) are unambiguously assignable to P. quica , exhibiting the craniodental traits of that species as diagnosed above.
The identity of Didelphys superciliaris Olfers, 1818 , which Patton and da Silva (1997, 2008) listed as a synonym of Philander frenatus , cannot now be determined. Both epithets were first published as nomina nuda by Illiger (1815), but their availability dates from Olfers (1818), who based his descriptions on material that he examined in Berlin (see appendix 5). However, whereas the application of frenatus can now be established based on an extant holotype, no type material of superciliaris is known to survive. Because the collections that Olfers examined in Berlin included specimens collected in both eastern Amazonia and southeastern Brazil, ( Voss and Angermann, 1997; see also appendix 5), the lost type of superciliaris could have come from either place: if from the former region, the name would be a junior synonym of P. opossum ; if from the latter, then it would be a senior synonym of P.quica . Because superciliaris has not, to our knowledge, been recognized as a valid taxon for almost two centuries, it would seem pointless to use this name to replace quica , the application of which is undisputed. Therefore, to fix the application of Didelphys superciliaris Olfers for the species that occurs in eastern Amazonia, we select as neotype a specimen in the American Museum of Natural History (AMNH 203348) consisting of the skin and skull of an adult male collected by personnel from the Instituto Oswaldo Cruz on 6 December 1960 near “Capim” (= São Domingos do Capim; 1°40´S, 47°47´W; Paynter and Traylor, 1991) at Km 92 on highway BR 14, Pará, Brazil.
HABITATS: Within the Atlantic Forest biome or ecoregion (Mata Atlântica), Philander quica seems to be eurytopic, occurring in a wide range of vegetation types including mature lowland rainforest (formerly the dominant climax vegetation of southeastern Brazil; Por, 1992), submontane forest, secondary growth, agricultural fields, and coastal restinga scrub ( Cerqueira et al., 1993; Bergallo, 1994; Bonvicino et al., 1997; Passamani et al., 2000; D’Andrea et al., 2007).
SPECIMENS EXAMINED (N = 66): Brazil — Espírito Santo, Engenheiro Reeve ( BMNH 3.9.4.110); Minas Gerais, Serra de Caparaó ( AMNH 8052–8053 About AMNH , 61851–61853 About AMNH ) ; Paraná, Parque Nacional do Iguaçu ( MVZ 197401 About MVZ ) , Roça Nova ( BMNH 3.7.1.108–3.7.1.110) ; Rio de Janeiro, Barreira ( ZMB 38069, 38072 View Materials , 38073 View Materials , 38076 View Materials , 38091 View Materials ) , Rio de Janeiro ( AMNH 133106 About AMNH , 133107 About AMNH ; ZMB 38063) , Sepetiba ( NMW 7687 View Materials /ST 1012) ; Rio Grande do Sul ( BMNH 84.2.8.29) ; Santa Catarina, Hansa ( BMNH 29.6.6.71), Jaraguá ( NMW B2529 View Materials ) , Teresópolis ( NMW 1671–1675 View Materials ) ; São Paulo, Boracéia ( MVZ 182777 About MVZ ; USNM 460503 About USNM ) , Fazenda Intervales ( MVZ 182066 About MVZ , 183246 About MVZ , 183247 About MVZ ) , Iguape ( USNM 542920 About USNM ) , Ilha de Sebastião ( MVZ 182067 About MVZ ) , Ilha do Cardoso ( FMNH 141589 About FMNH , 141590 About FMNH ) , São Sebastião ( BMNH 2.4.6.37–2.4.6.40), Ypanema ( NMW 2636 View Materials , 2638 View Materials , 2640 View Materials ) . Paraguay — Central , “Caroreni Viejo” (not located; ZMB 44285) ; Paraguarí, Sapucaí ( BMNH 2.11 .7.14, 2.11.7.48, 3.2.3.32–3.2.3.37 [type series of azaricus]; USNM 121412–121422, 121457, 121458), “Ipitimi” (= Ybytymí; ZMB 91277) ; San Pedro, Tacuatí ( USNM 293133 About USNM ) .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Philander quica ( Temminck, 1824 )
| Voss, Robert S., Díaz-Nieto, Juan F. & Jansa, Sharon A. 2018 |
Metachirus opossum azaricus
| Thomas, O. 1923: 604 |
Didelphis quica
| Hershkovitz, P. 1959: 342 |
| Temminck, C. J. 1824: 36 |