
Alpheus firmus Kim & Abele, 1988
|
publication ID |
https://doi.org/10.11646/zootaxa.5282.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:DF418763-8F0E-44DD-97C4-B123A81A8DB4 |
|
DOI |
https://doi.org/10.5281/zenodo.7921867 |
|
persistent identifier |
https://treatment.plazi.org/id/2A26026D-4B1D-FFEF-E7B8-F9B9FB1AFE6A |
|
treatment provided by |
Plazi (2023-05-09 07:04:28, last updated 2024-11-24 20:27:28) |
|
scientific name |
Alpheus firmus Kim & Abele, 1988 |
| status |
|
Alpheus firmus Kim & Abele, 1988 View in CoL
( Figs. 49 View FIGURE 49 , 50A, B View FIGURE 50 , 52F View FIGURE 52 )
Alpheus firmus Kim & Abele 1988: 93 View in CoL , fig. 39 (part.?); Abele & Kim 1989: 15; Wicksten & Hendrickx 1992: 4; Wicksten & Hendrickx 2003: 64 (part.?).
(?) Alpheus firmus View in CoL .— Alvarez et al. 1996: 716, fig. 2 [= A. cf. firmus Kim & Abele, 1988 View in CoL ; see below].
Material examined. Panama: 1 male (cl 6.9 mm), MNHN-IU-2018-5574, Veraguas, Coiba Archipelago, Coiba, Bahía Santa Cruz , intertidal mudflat with rocks, leg. A. Anker et al., 21.03.2007 [AA-07-145]; 1 male (cl 11.0 mm), 1 ov. female (cl 12.8 mm), USNM 237798 About USNM , Panama Canal, Pacific side entrance, Miraflores Locks , NMNH-STRI sta. 203, 26.08.1974; 1 male, 1 female (cl indet., specimens in poor condition, not deposited, see below) , Panama Canal, Pacific side entrance, Miraflores Locks, leg. J. Jara et al., 21.07.2005 [AA-05-189] .
Tentative identification. Alpheus cf. firmus . Panama: 1 male (cl 10.0 mm), MNHN-IU-2018-5577, Veraguas, Puerto Vidal, mangrove creek, in mud under rocks at low tide, leg. A. Anker, 19.03.2007 [AA-07-092B]; 1 ov. female (cl 10.5 mm), MNHN-IU-2018-5576, same collection data as for previous specimen [AA-07-092A]; 1 male (cl 9.3 mm), MNHN-IU-2018-5575, same collection data as for previous specimens [AA-07-091].
Description. See Kim & Abele (1988) for detailed description and illustrations.
Colour pattern. Type A colour morph ( Alpheus firmus sensu Kim & Abele 1988 ): body uniform purplish to pinkish brown; posterior edges of pleonites and rostrum darker reddish-brown; antennular and antennal flagella greenish yellow; major and minor chelae greenish to olive-brown with paler green-yellow areas, fingers distally pinkish; walking legs reddish; uropod with bluish tinge distally ( Figs. 49 View FIGURE 49 , 50A, B View FIGURE 50 ). Type B colour morph ( Alpheus cf. firmus ): greenish to olive-brown with whitish saddle on carapace and transverse whitish areas on pleon resulting in banded pattern; antennular and antennal flagella greenish yellow; major and minor chelae green-brown with paler greenish areas, fingers distally reddish; walking legs reddish; uropod with deep blue and brick-red areas distally ( Fig. 50D–F View FIGURE 50 ). See discussion of the two colour morphs below.
Type locality. Miraflores Locks , Pacific entrance to Panama Canal, Panama.
Distribution. Tropical eastern Pacific: Panama (Punta Paitilla, Panama Canal, Puerto Vidal, Coiba Island) to Mexico (Nayarit) ( Fig. 52F View FIGURE 52 ) ( Kim & Abele 1988; Alvarez et al. 1996; present study); however, A. firmus sensu Kim & Abele (1988) may be restricted to Panama (see discussion below).
Common name proposed. Panama Canal snapping shrimp.
Ecology and biology. Alpheus firmus is confined to mudflats and mangroves with abundant wood debris and rocks, living in self-made burrows, sometimes dug under large rocks or decaying logs, mainly in the intertidal and adjacent shallow subtidal areas (a few meters in depth). It appears to be a euryhaline species, capable of tolerating brackish conditions. The large number of eggs and their small size suggest an extended larval development.
Taxonomic remarks. The type series of A. firmus (USNM) is composed of 82 adult specimens collected at Miraflores Locks ( Panama Canal) and Punta Paitilla, Panama City. In July 2005, a male-female pair of A. firmus ( Fig. 49 View FIGURE 49 ) was collected at the type locality of the species and kept in the running sea water tanks of the Naos Laboratories of the Smithsonian Tropical Research Institite (STRI). Unfortunately, due to an accidental rupture of sea water supply, both shrimps died and were found partly disintegrated; therefore, they were preserved in ethanol, but not deposited in a museum collection due to their poor condition. Another specimen, a smaller male, was collected in 2007 on a mudflat of Santa Cruz Bay, Coiba Island ( Fig. 50A–C View FIGURE 50 ). These three specimens correspond well to the description and illustrations of the holotype of A. firmus in Kim & Abele (1988).
Kim & Abele (1988) noted some variation in the presence / absence of a small, sharp tooth on the antennal basicerite, as well as in the relative length ratio of the penultimate and ultimate articles of the third maxilliped in the Panamanian material of A. firmus . The variation in these two features seems to be correlated: all specimens with a tooth on the basicerite also had a relatively shorter penultimate article of the third maxilliped (ratio ultimate vs. penultimate article 1.3–1.6), compared to the specimens with an unarmed basicerite (ratio ultimate vs. penultimate article 1.2). For instance, in the illustrated holotype of A. firmus , the basicerite has a small tooth and the ratio between the ultimate and penultimate articles is approximately 1.3 ( Kim & Abele 1988: fig. 39d). In the specimen of A. firmus from Coiba, Panama, the basicerite is armed with a small tooth and the ratio between the ultimate and penultimate articles is about 1.5, whereas in the specimens tentatively identified as A. cf. firmus from Puero Vidal, Panama, the basicerite is unarmed and the ratio between the ultimate and penultimate articles is at most 1.2, thus confirming observations by Kim & Abele (1988). Alvarez et al. (1996), reporting A. firmus from Nayarit, Mexico, noted that their specimens also were lacking a tooth on the antennal basicerite and had a long ultimate article of the third maxilliped (twice as long as the penultimate instead of slightly longer, as in the type material). However, in the figure of the third maxilliped provided by these authors, the ultimate article is only ~1.6 times longer than the penultimate article ( Alvarez et al. 1996: fig. 2b), although both articles are noticeably broader than their counterparts in the holotype ( Kim & Abele 1988: fig. 39d). In addition, the colour pattern of the specimens from Miraflores Locks and Coiba Island identified as A. firmus (type A, with a non-banded pleon, Figs. 49 View FIGURE 49 , 50A, B View FIGURE 50 ), is somewhat different from that of the specimens assigned to A. cf. firmus (type B, with a banded pleon, Fig. 50D–F View FIGURE 50 ).
The variation in both morphology (presence vs. absence of a tooth on the antennal basicerite, ratio and proportions of the penultimate and ultimate articles of the third maxilliped) and colour pattern (non-banded vs. banded pleon) in the Panamanian material and the presence of only morphotype in the Mexican material of A. firmus ( Alvarez et al. 1996) , suggests that a second species may be present in the tropical eastern Pacific, ranging from Mexico ( Alvarez et al. 1996) to Panama ( Kim & Abele 1988; present study), where it overlaps with the distributional range of A. firmus . This taxonomic problem will be addressed by the author in the near future, after a careful examination of the entire type series of A. firmus in the USNM (possible only after the end of the Covid-19 pandemic), DNA analyses of the Panamanian material collected in 2007, and a thorough morphological and genetic comparison of A. cf. firmus with A. zarenkovi Anker & Pachelle, 2015 , described from the same geographical area ( Anker & Pachelle 2015). For the time being, it seems more prudent to treat A. cf. firmus separately from A. firmus and to restrict the distributional range of the latter species to Panama (as shown in Fig. 52F View FIGURE 52 ).
Alpheus firmus appears to be most closely related to the amphi-Atlantic Atlantic A. pontederiae and several members of the transisthmian A. heterochaelis species complex (possibly non-monophyletic grouping), which includes the western Atlantic A. heterochaelis Say, 1818 , A. buckupi Almeida, Terossi, Araújo-Silva & Mantelatto, 2013 and A. petronioi Almeida, Terossi & Mantelatto, 2014 , and the eastern Pacific A. distinctus Kim & Abele, 1988 , all characterised by the presence of a well-developed balaeniceps condition on the male minor chela. However, A. firmus (sensu Kim & Abele 1988) can be separated from all these species by the longer and relatively slenderer penultimate article of the third maxilliped ( Kim & Abele 1988: fig. 39d); specifically from A. pontederiae by the absence of a distinct mesial subdistal ridge on the major chela pollex (idem: fig. 39f; cf. Fig. 47D View FIGURE 47 , 48C View FIGURE 48 ; Christoffersen 1984: fig. 4a) and the third and fourth pereiopods with a trigonal, feebly broadened dactylus (idem: fig. 39k; cf. Christoffersen 1984: fig. 4h); from A. heterochaelis by the distinctly shorter rostrum and the more gently sloping dorsal shoulder of the major chela (idem: fig. 39b, f; cf. Williams 1984: fig. 65a, b); from A. buckupi by the much shorter rostrum, the less marked rostral carina, the less concave lateral margin of the antennal scaphocerite, and the distomesially unarmed cheliped merus (idem: fig. 39b, f; cf. Almeida et al. 2013: figs. 1A, 2A); from A. petronioi by the shorter rostrum and less marked rostral carina, and the longer second article of the antennular pedunce (idem: fig. 39b; cf. Almeida et al. 2014: fig. 1B); and from A. distinctus by the more gently sloping dorsal shoulder of the major chela, the much less sculptured male minor chela, and the different ratio of the first and second carpal subarticle of the second pereiopod (idem: fig. 39f, h, j, 40f, h, i). Alpheus firmus also has affinities to some Indo-West Pacific species of the A. euphrosyne — A. microrhynchus complex, but can be easily separated from all of them by the elongated penultimate article of the third maxilliped, in combination with other morphological features (e.g., trigonal dactylus of the walking legs, major chela pollex without mesial subdistal ridge, weak sculpture of the minor chela, several features of the rostro-orbital region, antennules, antennae, etc.) and colour pattern.
Abele, L. G. & Kim, W. (1989) The decapod crustaceans of the Panama Canal. Smithsonian Contributions to Zoology, 482, 1 - 50. https: // doi. org / 10.5479 / si. 00810282.482
Almeida A. O. de, Terossi M., Araujo-Silva C. L. & Mantelatto, F. L. (2013) Description of Alpheus buckupi spec. nov., a new amphi-Atlantic snapping shrimp (Caridea: Alpheidae), based on morphological and molecular data. Zootaxa, 3652 (4), 437 - 452. https: // doi. org / 10.11646 / zootaxa. 3652.4.3
Almeida, A. O. de, Terossi, M. & Mantelatto, F. L. (2014) Morphology and DNA analyses reveal a new cryptic snapping shrimp of the Alpheus heterochaelis Say, 1818 (Decapoda: Alpheidae) species complex from the western Atlantic. Zoosystema, 36, 53 - 71. https: // doi. org / 10.5252 / z 2014 n 1 a 4
Alvarez, F., Camacho, M. E. & Villalobos, J. L. (1996) The first species of Prionalpheus from the eastern Pacific, and new records of caridean shrimp (Crustacea: Decapoda: Caridea) from the western coast of Mexico. Proceedings of the Biological Society of Washington, 109, 715 - 724.
Anker, A. & Pachelle, P. P. G. (2015) Two new snapping shrimps (Decapoda, Caridea, Alpheidae, Alpheus) from the tropical eastern Pacific. Arthropoda Selecta, 24, 247 - 258.
Christoffersen, M. L. (1984) The western Atlantic snapping shrimps related to Alpheus heterochaelis Say (Crustacea, Caridea) with the description of a new species. Papeis Avulsos de Zoologia, 35, 189 - 208.
Kim, W. & Abele, L. G. (1988) The snapping shrimp genus Alpheus from the eastern Pacific (Decapoda: Caridea: Alpheidae). Smithsonian Contributions to Zoology, 454, 1 - 119. https: // doi. org / 10.5479 / si. 00810282.454
Say, T. (1818) An account of the Crustacea of the United States, part 5. Journal of the Academy of Natural Sciences at Philadelphia, 1, 235 - 253.
Wicksten, M. K. & Hendrickx, M. E. (1992) Checklist of penaeoid and caridean shrimps (Decapoda: Penaeoidea, Caridea) from the eastern tropical Pacific. Proceedings of San Diego Society of Natural History, 9, 1 - 11.
Wicksten, M. K. & Hendrickx, M. E. (2003) An updated checklist of benthic marine and brackish water shrimps (Decapoda: Penaeoidea, Stenopodidea, Caridea) from the eastern tropical Pacific. In: Hendrickx, M. E. (Ed.), Contribuciones al estudio de los crustaceos del Pacifico Este. Instituto de Ciencias del Mar y Limnologia, Universidad Nacional Autonoma de Mexico, Mexico City, pp. 49 - 76.
Williams, A. B. (1984) Shrimps, lobsters, and crabs of the Atlantic coast of the eastern United States, Maine to Florida. Smithsonian Institution Press, Washington, D. C., 550 pp.
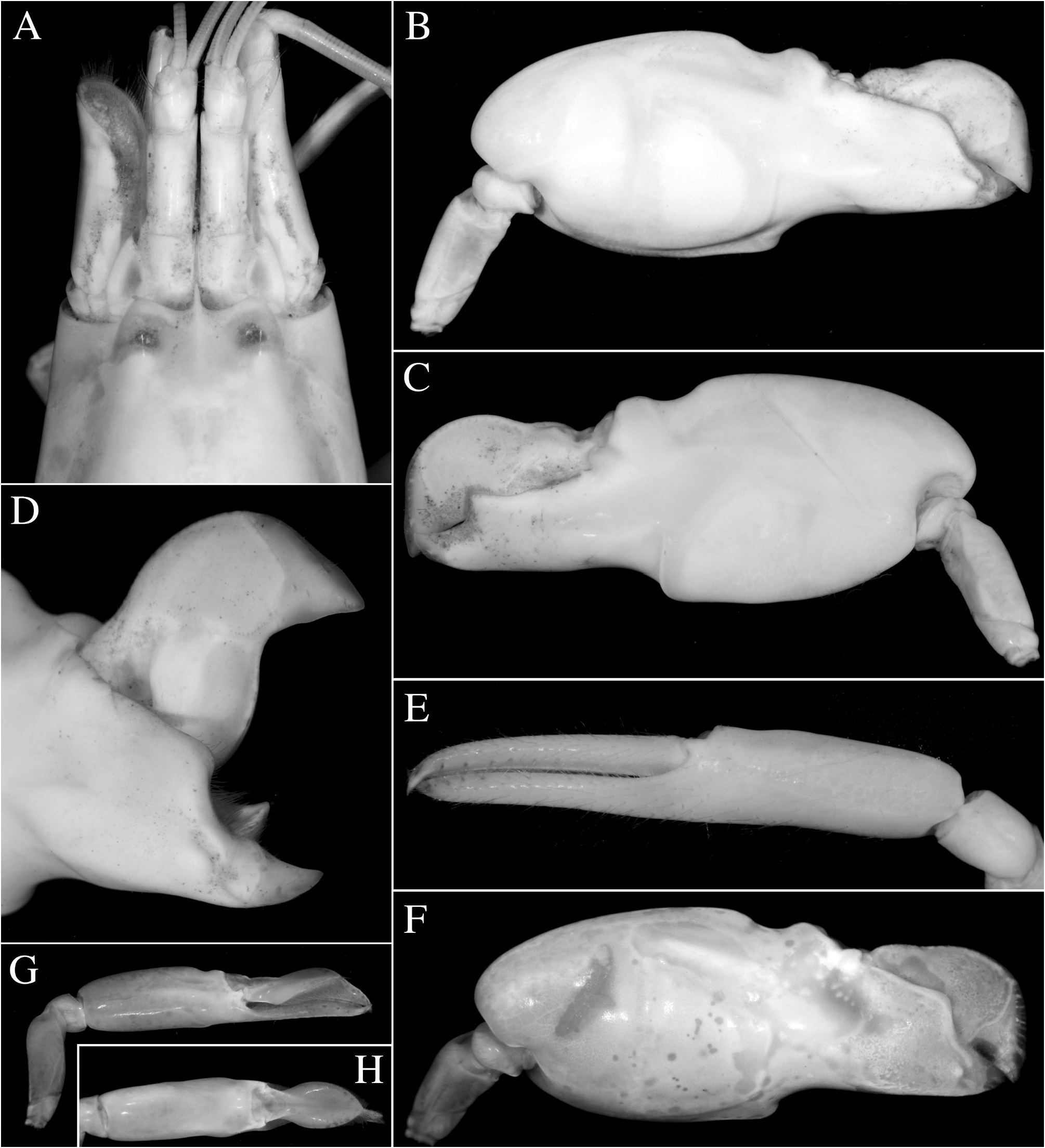
FIGURE 47. Alpheus pontederiae de Rochebrune, 1883: female (cl indet.) from Orinoco Delta, Venezuela (MZUSP) [A–E]; male (cl indet.) from the same locality (MZUSP) [F–H]; A—frontal region, dorsal view; B, F—major (left) cheliped, mesial view; C—same, lateral view; D—same, chela fingers opened, mesial view; E—minor (right) cheliped, mesial view; G—minor (right) cheliped, lateral view; H, same, chela, dorsal view.

FIGURE 48. Alpheus pontederiae de Rochebrune, 1883: male (cl indet.) from near Bragança, Pará, Brazil (MPEG) [A–C]; male (cl ~6.0 mm) from S„o Tomé, S„o Tomé and Príncipe (OUMNH.ZC. 2011.06.8) [D] and its habitat near Porto Alegre, S„o Tomé [E]; A, D—habitus, dorsal view; B—same, lateral view; C—frontal region and chelipeds, dorsal view; E—mangrove channel near Porto Alegre, S„o Tomé, collection locality of the specimen in D. Photographs by the author.

FIGURE 49. Alpheus firmus Kim &Abele, 1988: female (cl indet.) [A] and male (cl indet.) [B] from Miraflores Locks, Panama Canal, Panama (specimens not deposited, see text); A, B—habitus, dorsal view. Photographs by the author.

FIGURE 50. Alpheus firmus Kim & Abele, 1988: male (cl 6.9 mm) from Coiba, Panama (MNHN-IU-2018-5574) [A–C]; A, B—habitus, dorsal view; C—habitat, estuarine mangrove in Bahía Santa Cruz, Coiba. Alpheus cf. firmus Kim & Abele, 1988: male (cl 10.0 mm) from Puerto Vidal, Panama (MNHN-IU-2018-5577) [D–F]; D, E—habitus, dorsal view; F—same, lateral view. Photographs by the author.
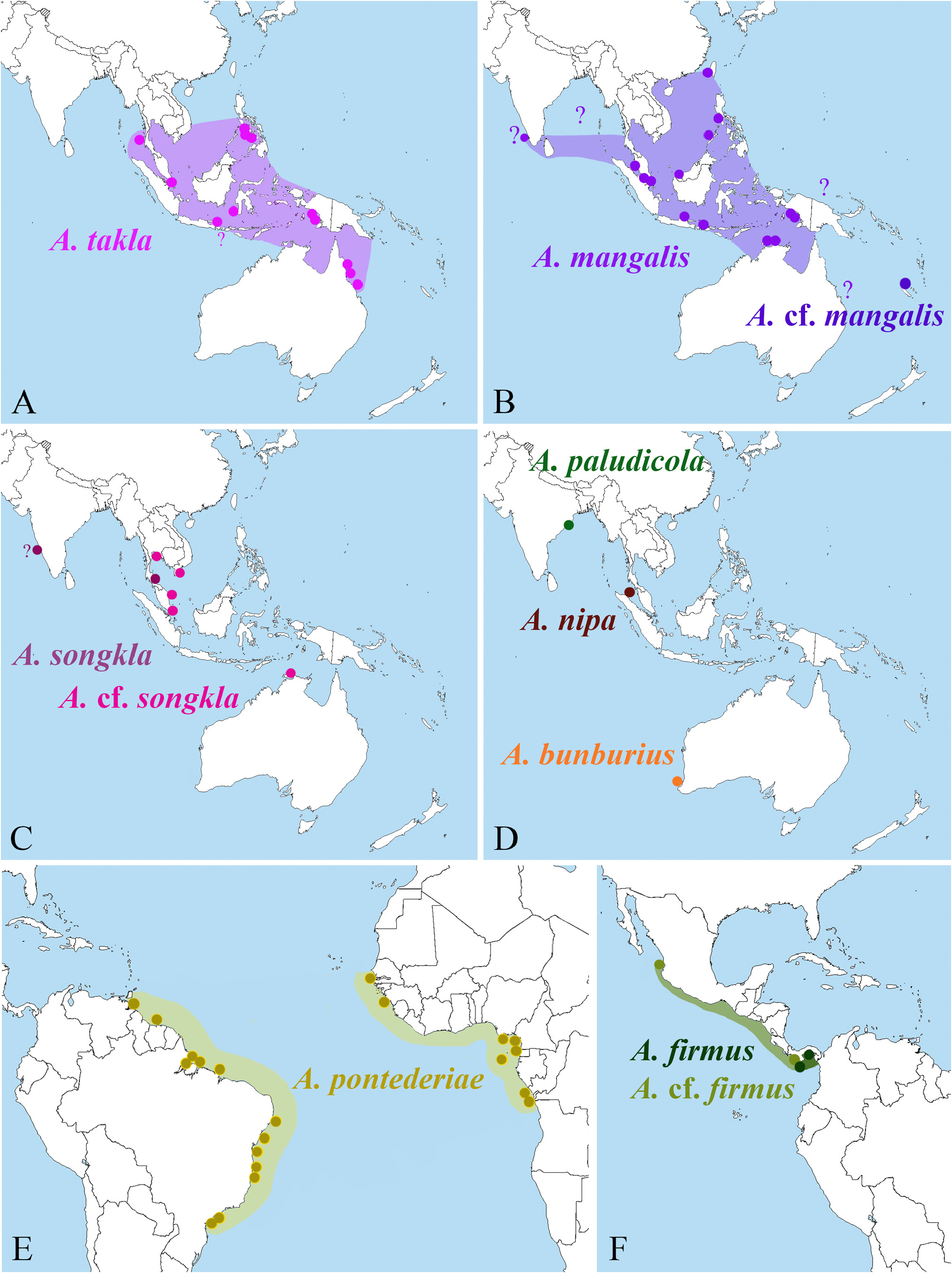
FIGURE 52. Distributional maps for species of Alpheus treated in this study:A—A. takla sp. nov.; B—A. mangalis sp. nov. and A. cf. mangalis sp. nov.; C—A. songkla Banner & Banner, 1966 and A. cf. songkla (see text); D—A. paludicola Kemp, 1915, A. nipa Banner & Banner, 1985 and A. bunburius Banner & Banner, 1982; E—A. pontederiae de Rochebrune, 1883; F—A. firmus Kim & Abele, 1988 and A. cf. firmus (see text).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alpheus firmus Kim & Abele, 1988
| Anker, Arthur 2023 |
Alpheus firmus Kim & Abele 1988: 93
| Wicksten, M. K. & Hendrickx, M. E. 2003: 64 |
| Wicksten, M. K. & Hendrickx, M. E. 1992: 4 |
| Abele, L. G. & Kim, W. 1989: 15 |
| Kim, W. & Abele, L. G. 1988: 93 |