
Squalus bahiensis, De, Sarah T., De, Marcelo R. & Gomes, Ulisses L., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4133.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E7276A63-67C8-4BC5-8419-2EBDAE4432B0 |
|
DOI |
https://doi.org/10.5281/zenodo.6075730 |
|
persistent identifier |
https://treatment.plazi.org/id/282F878E-FFA5-FFB6-14C0-2171FD565F68 |
|
treatment provided by |
Plazi |
|
scientific name |
Squalus bahiensis |
| status |
sp. nov. |
Squalus bahiensis View in CoL sp. nov.
( Figs. 26–33 View FIGURE 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 View FIGURE 33 ; Tables 7–8 View TABLE 7 View TABLE 8 )
Suggested common names: Northeastern Brazilian dogfish; Cação-bagre-da-Bahia (Portuguese).
Squalus fernandinus View in CoL (not Molina): Fowler, 1936: 71 (description; South Atlantic); Bigelow & Schroeder, 1948: 480 (revision; Brazil).
Squalus cubensis View in CoL (not Howell-Rivero): Kondyurin & Myagkov, 1984: 118 –120, fig. 1B (revision; Western Atlantic); Nunan & Senna, 2007: 169, 170 (cited; Brazil).
Squalus blainvillei View in CoL (not Risso):? Lucena & Lucena, 1981: 2, 4, 5, fig. 3 (listed; Brazil);? Menni et al., 1984: 84 (listed; Argentina, Uruguay);?Canizarro et al., 1994: 113 (cited; Western Atlantic Ocean);?Nion et al., 2002: 4 (listed; Uruguay);? Meneses & Paesch, 2003: 8, 25 (cited; Argentina, Uruguay);?Saéz et al., 2010: 623 (identification key; Chile); Viana, 2011 (in part): 57–91, figs. 18–34 (revision; Brazil).
Squalus mitsukurii View in CoL (not Jordan & Snyder): Calderón, 1994: fig. 5B (cited; Brazil);? Lessa et al., 1999: 61, 150 (cited, listed; Northeast Brazil); Compagno, 2002: 385 (description; Northeast Brazil); Jablonski et al., 2006: 177 (cited; Brazil); Louro & Rossi-Wongtschowski, 2007: 18, 27, 28, 30, 49 (cited; Brazil);?Fischer et al., 2006: 495–501 (cited; Northeastern Brazil).
Squalus View in CoL sp. of the blainvillei View in CoL group: Gomes et al., 1997: 93 –95, 109 (listed; Brazil); Marques, 1999 (cited; Brazil).
Squalus View in CoL of the blainvillei View in CoL / mitsukurii View in CoL group: Gadig, 2001 (in part): 29, 36, 54, 55, 57, 58, fig. 29 (listed; Brazil).
Squalus View in CoL sp. B: Soto, 2001: 96 (listed; Brazil); Soto & Mincarone, 2004: 79 –82 (listed; Brazil).
Squalus View in CoL sp. 1: Gomes et al., 2010: 44, 45 (cited; Brazil).
Holotype. MNRJ 30180, adult male, 590 mm TL, Bahia coast, Brazil, 1548'30''S, 3835'16''W, 599 m. Collected on 12 June 2000, Station 0 510, Thalassa cruise, Revizee program.
Paratypes (2 specimens). MNRJ 30178, adult male, 615 mm TL, 599 m; MNRJ 30179, adult male, 690 mm TL, 599 m. Same locality as holotype.
Diagnosis. Squalus bahiensis sp. nov. is distinct from all species of the S. megalops group by having pectoral fins with free rear tips rounded (vs. free rear tips pointed), snout blunt and elongate (vs. snout rounded and short), and dermal denticles tricuspid and rhomboid (vs. dermal denticles unicuspid and lanceolate). Squalus bahiensis sp. nov. is distinct from S. mitsukurii by having a narrower interorbital space (8.2%, 7.9%–8.3% TL vs. 9.3%, 9.1%– 9.8% TL in S. mitsukurii ), and shorter lower caudal fin lobe (preventral caudal margin 11.4%, 10.9%–11.3% TL vs. 12.1%, 11.5%–12.4% TL). Squalus bahiensis sp. nov. can be easily distinguished from S. blainvillei from the Mediterranean Sea by having lower dorsal fins (first dorsal-fin height 6.9%, 7.0%–7.1% TL vs. 8.2%, 7.3%–8.7% TL in S. blainvillei ; second dorsal-fin height 5.3%, 4.2%–4.4% TL vs. 7.2%, 6.6%–7.5% TL in S. blainvillei ), and a shorter upper labial furrow (its length 2.3%, 2.2% TL vs. 2.5%, 2.4%–2.7% TL in S. blainvillei ). Squalus bahiensis sp. nov. is distinct from S. lobularis by: caudal fin somewhat slender vs. upper caudal lobe rectangular in S. lobularis ; second dorsal-fin spine not reaching dorsal-fin apex vs. reaching dorsal-fin apex in S. lobularis ; shorter pectoral-fin inner margin (its length 8.1%, 8.4%–8.9% TL vs. 10.5%, 9.2%–11.0% TL in S. lobularis ); and clasper outer length greater (5.0%, 4.6%–5.1% TL vs. 4.3% TL in S. lobularis ).
Description. External morphology. Measurements and meristic data are summarized in Tables 7–8 View TABLE 7 View TABLE 8 . Body slender and fusiform, arched dorsally and somewhat flattened ventrally; its greatest depth at head and abdomen (head height 9.3%, 9.6%–10.0% TL; abdomen height 10.1%, 9.6%–10.5% TL) with head height corresponding to 0.9 (1.0–1.0) times trunk height and 0.9 (1.1–1.1) times abdomen height. Head rather elongate (length 21.4%, 22.8%–22.9% TL), narrower at nostrils (width 7.0%, 7.0%–7.1% TL), and much broader at gills (11.9%, 12.4%– 12.6% TL). Snout markedly blunt and broad at tip (not obtuse) and elongate (preorbital length 7.3%, 7.4%–7.9% TL); anterior margin of nostrils bilobate, positioned laterally and nearer to the mouth than to snout tip (prenarial length 1.1, 1.0–1.2 times inner nostril-labial length); prenarial length 1.4 (1.4–1.4) times eye length; internarial space 1.2 (1.3–1.4) times eye length. Eyes oval and horizontal, placed nearer to snout tip than to gill slits; its anterior margin convex and posterior margin notched; eyes large, their length 2.0 (1.9–2.7) times its height. Prespiracular length 12.2% (12.2%–12.9% TL), corresponding to 0.6 (0.5–0.6) times prepectoral length. Spiracles crescent-shaped, located more dorsally behind eyes; spiracle length 0.4 (0.3–0.5) times eye length. Prebranchial length 17.9% (18.4%–18.8% TL), corresponding to 1.5 (1.4–1.5) times prespiracular length. Gill slits vertical and low, fifth gill slit height 1.0 (1.1–1.2) times greater than first gill slit height; gill slits just anterior to pectoral fins.
Preoral length 9.9% (10.2%–10.5% TL), corresponding to at least 1.4 times mouth width. Mouth arched and somewhat broad, its width 1.7 (1.5–1.6) times internarial space and 1.4 (1.4–1.5) times prenarial length; upper labial furrow long, its length 2.3% (2.2%–2.2% TL) with a slender fold; lower labial furrow also elongate, although without a fold. Teeth unicuspid, similar in both jaws, broad and compressed labial-lingually at crown, imbricate laterally; upper teeth smaller and narrower than lower teeth; cusp small and heavy, markedly oblique and upturned, directed laterally; mesial cutting edge conspicuously convex; mesial heel notched; distal heel strongly rounded; apron thick; two series of functional teeth on upper jaws of holotype and three series in paratypes; two series of functional teeth in lower jaws; tooth rows varying from 13–14 (13–14 paratypes) in upper jaw and 11–12 (11–11 paratypes) in lower jaw ( Fig. 28 View FIGURE 28 ).
Pre-first dorsal fin length 29.9% (29.7%–30.9% TL), corresponding to 1.4 (1.3–1.4) times prepectoral length. Origin of first dorsal fin preceding pectoral free rear tips. First dorsal fin wide at fin web and rather large ( Fig. 29 View FIGURE 29 ), its length 12.8% (13.8%–13.8% TL), corresponding to 1.8 (1.9–2.0) times its height; anterior margin convex and elongate, its length 10.5% (10.3%–10.6% TL); posterior margin markedly concave and large, its length 7.9% (9.2%–9.6% TL); first dorsal-fin apex strongly rounded and slender; first dorsal-fin free rear tip triangular; first dorsal-fin inner margin length 5.7% (6.4%–6.9% TL); first dorsal fin low, its height 6.9% (7.0%–7.1% TL) and corresponding to 1.2 (1.0–1.1) times its inner margin length, and 0.9 times preorbital length. First dorsal-fin spine slender and small, its length 2.8% (2.9%–3.0% TL) and corresponding to 0.4 times first dorsal-fin height, not reaching first dorsal-fin apex. First dorsal-fin length 1.1 times second dorsal-fin length. Interdorsal space 1.2 (1.1– 1.2) times prepectoral length and 2.4 (2.0–2.4) times dorsal-caudal space. Pre-second dorsal fin length 2.8 times prepectoral length and 4.3 (4.2–4.4) times pectoral anterior margin length. Second dorsal fin slender, small, its length 11.6% (12.1%–12.6% TL) and 2.2 (2.8–2.9) times its height; anterior margin convex and short, its length 9.4% (9.3%–9.7% TL); posterior margin conspicuously concave and falcate, its length 5.0% (5.7%–5.7% TL); second dorsal-fin apex rounded and lobe-like; second dorsal-fin free rear tip triangular; second dorsal-fin inner margin somewhat elongate, its length 5.3% (5.6%–5.6% TL); second dorsal fin also low, its height 1.0 (0.8–0.8) times its inner margin length. Second dorsal-fin spine thin and short, its length 0.8 (0.8–1.0) times second dorsalfin height and 1.5 (1.2–1.5) times greater than first dorsal-fin spine; second dorsal-fin spine not reaching second dorsal-fin apex ( Fig. 29 View FIGURE 29 ).
Prepectoral length 21.7% (22.0%–22.8% TL). Pectoral fins markedly narrow and small (anterior margin length 14.3%, 14.4%–14.5% TL); anterior margin 1.8 (1.6–1.7) times greater in length than inner margin length and 1.4 (1.3–1.4) times posterior margin length; pectoral-fin anterior margin straight and posterior margin convex; inner margin markedly concave, its length 8.1% (8.4%–8.9% TL); pectoral-fin apex rounded and lobe-like; pectoral-fin free rear tips also rounded and lobe-like, reaching the horizontal line traced from pectoral fin apex ( Fig. 27 View FIGURE 27 ).
Prepelvic length 44.4% (45.5%–48.6% TL). Pectoral-pelvic space 0.7 (0.8–0.9) times pelvic-caudal space. Pelvic fins also narrow, its anterior and posterior margins straight; pelvic-fin free rear tips slightly pointed, triangular and elongate, its length 5.8% (5.1%–5.4% TL). Origin of pelvic fins 2.8 (2.9–3.0) times the distance between the origins of the two dorsal fins, closer to the first dorsal fin than to second dorsal fin in holotype (somewhat closer to midline between the origins of two dorsal fins in paratypes); pectoral-pelvic space short, its length 19.7% (19.5%–20.3% TL) and corresponding to 0.7 (0.7–0.8) times pelvic-caudal space. Claspers cylindrical and slender, compressed dorsoventrally; claspers large, extending far posterior to pelvic-fin free rear tips, its outer length among 5.0% (4.6%–5.1% TL); clasper inner margin 1.3 (1.3–1.4) times greater in length than pelvic-fin inner margin; clasper groove longitudinal and very elongate, positioned dorsally; apopyle very narrow, located anteriorly in clasper groove; hypopyle also narrow, located posteriorly in clasper groove; rhipidion markedly large, blade-like and slender, placed medially at distal end of clasper ( Fig. 30 View FIGURE 30 ).
Caudal peduncle with soft lateral caudal keels from opposite second dorsal-fin free rear tip to caudal fin origin; upper and lower caudal pits profound. Caudal fin with straight dorsal caudal margin, its length 21.9% (20.0%– 21.4% TL) and corresponding to 1.0 (0.9 in paratypes) times head length and 1.9 (1.8–1.9) times preventral caudal margin length; upper postventral caudal margins strongly convex ( Fig. 31 View FIGURE 31 ); lower postventral caudal margin convex; posterior caudal tip rounded; preventral caudal margin straight and short, its length 2.0 times (2.1 in paratypes) pelvic-fin inner margin length; ventral caudal tip also rounded; caudal fin narrow at lobes and conspicuously concave at caudal fork, its width 6.7% (6.4%–7.1% TL).
Dermal denticles ( Fig. 32 View FIGURE 32 ). Description based on paratype MNRJ 30178. Denticles tricuspid and imbricate dermal denticles, very broad at crown, their length somewhat equal to their width; cusps triangular and wide with median cusp larger than lateral cusps; lateral cusps forming a right angle with median cusp; median anterior projection conspicuous and rounded with small posterolateral expansions on each side; median ridge conspicuous and bifurcate, forming a superficial furrow anteriorly, reaching the median cusp tip; two lateral ridges on each side of the denticle, thick and prominent, reaching the lateral cusp tips; lateralmost ridge often shorter than the medial ridge. In holotype, dermal denticles more slender and not imbricate; cusps very thin and cylindrical; lateral cusps forming a prominent concavity with the median cusp on each side.
Coloration ( Fig. 26 View FIGURE 26 ). Body gray dorsally, white ventrally and on its posterior half. First dorsal fin gray, darker at apex and whitish at base; anterior margin slightly white on first half; posterior margin white. Second dorsal fin also gray and darker at apex; posterior margin slightly white. First and second dorsal-fin spines white, slightly gray anteriorly. Pectoral fins gray dorsal and ventrally, whitish at ventral base; pectoral-fin posterior margin uniformly white. Pelvic fins light gray dorsally and ventrally; pelvic-fin posterior margin white. Caudal fin dark gray, whitish over vertebral column; dorsal caudal margin white; upper and lower postventral caudal margins whitish; posterior caudal tip broadly white; ventral caudal tip also white; preventral caudal margin somewhat whitish; caudal stripe small and gray.
Vertebral counts ( Table 8 View TABLE 8 ). Monospondylous vertebrae 43 in holotype (43–45 in paratypes); precaudal vertebrae 87 (87); total vertebrae 117 (115–116).
Geographical distribution ( Fig. 33 View FIGURE 33 ). Squalus bahiensis is apparently endemic to the coast near Salvador, Bahia ( Brazil).
Etymology. The epithet is in reference to the state of Bahia, northeastern Brazil, from where this species is described.
Remarks. Squalus mitsukurii and the Mediterranean species S. blainvillei ( Risso, 1826) are often listed in the literature from the northern Brazilian coast to southern Argentina (e.g. Miranda Ribeiro, 1907; Compagno, 1984; Calderón, 1994; Compagno et al., 2005; Fischer et al., 2006; Viana, 2011). These two species were probably misidentified in these regions for sharing characters of species of the S. mitsukurii group, such as an elongate snout, tricuspidate dermal denticles, and low dorsal fins, which are also present in both S. bahiensis and S. lobularis . Squalus bahiensis , however, can be easily distinguished from all species of this group by the absence of a black caudal blotch in the caudal fin. Squalus bahiensis further differs from S. lobularis (its most similar congener) by a shorter first dorsal-fin anterior margin (its length in S. bahiensis 10.5%, 10.3%–10.6% TL vs. 11.1%, 10.3%–11.9% TL in S. lobularis ), and greater interdorsal space (26.5%, 24.4%–26.8% TL vs. 24.6%, 21.9%–24.1% TL, respectively).
Squalus bahiensis can also be distinguished from the Japanese S. mitsukurii by a more elongate precaudal length (79.5%, 79.2%–81.2% TL vs. 77.5%, 76.9%–78.6% TL in S. mitsukurii ), pre-second dorsal fin length (61.5%, 61.0%–63.8% TL vs. 61.0%, 58.4%–60.5% TL in S. mitsukurii ), and first gill slit closer to snout tip in S. bahiensis than in S. mitsukurii (prebranchial length 17.9%, 18.4%–18.8% TL vs. 20.4%, 19.9%–20.0% TL in S. mitsukurii ). Squalus bahiensis has a body depth almost equal from the head to abdomen (head height 0.9, 1.1–1.1 times trunk height, and 0.9, 1.0–1.0 times abdomen height) while S. mitsukurii has its greatest depth in the head (head height 1.2, 1.0–1.4 times trunk height and 1.6, 1.2–1.7 times abdomen height). It is also easily distinct from S. montalbani by having a more slender and deep body (vs. markedly robust body in S. montalbani ), and a shorter ventral caudal lobe (its length 11.4%, 10.9%–11.3% TL vs. 11.5%–13.2% TL in S. montalbani ).
Squa'us Squa'us Hο Hο MNRJ MΖUSP
bahiensis sp. nov. quasimodo sp. nov. Measurements 1οtype Paratypes 1οtype Paratypes 30180 N Range x SD 118707 N Range x SD ΤL (mm) 585.0 2 615.0 - 690.0 652.5 53.0 700.0 3 660.0 - 830.0 745.0 85.0 PCL 79.5 2 79.2 - 81.2 80.2 1.4 82.9 3 78.5 - 80.1 79.4 0.8 PD2 61.5 2 61.0 - 63.8 62.4 2.0 63.6 3 60.1 - 62.3 61.5 1.2 PD1 29.9 2 29.7 - 30.9 30.3 0.8 32.9 3 28.4 - 31.8 30.3 1.7 SvL 47.0 2 47.2 - 50.7 48.9 2.5 49.7 3 47.0 - 50.6 48.4 1.9 PP2 44.4 2 45.5 - 48.6 47.0 2.1 49.3 3 44.7 - 48.0 46.2 1.6 PP1 21.7 2 22.0 - 22.8 22.4 0.6 23.7 3 21.8 - 22.6 22.2 0.4 HDL 21.4 2 22.8 - 22.9 22.8 0.1 24.7 3 22.2 - 23.0 22.6 0.4 PG1 17.9 2 18.4 - 18.8 18.6 0.3 20.7 3 17.4 - 19.2 18.5 1.0 PSP 12.2 2 12.2 - 12.9 12.6 0.5 12.8 3 11.5 - 12.6 12.2 0.6 PΟB 7.3 2 7.4 - 7.9 7.7 0.3 7.9 3 6.8 - 7.7 7.4 0.5 PRN 5.0 2 4.9 - 5.1 5.0 0.1 4.8 3 4.5 - 4.9 4.7 0.2 PΟR 9.9 2 10.2 - 10.5 10.3 0.2 10.2 3 9.5 - 10.0 9.7 0.3 INLF 4.6 2 4.2 - 4.9 4.6 0.5 4.7 3 4.0 - 4.7 4.5 0.4 MΟW 7.2 2 7.2 - 7.7 7.5 0.3 7.8 3 7.3 - 7.8 7.5 0.2 ULA 2.3 2 2.2 - 2.2 2.2 0.0 2.6 3 2.2 - 2.5 2.4 0.1 INW 4.2 2 4.7 - 4.8 4.8 0.1 5.0 3 3.9 - 4.1 4.0 0.1 INΟ 8.2 2 7.9 - 8.3 8.1 0.3 8.8 3 7.7 - 8.0 7.9 0.2 EΥL 3.6 2 3.5 - 3.8 3.6 0.2 3.5 3 3.1 - 4.9 4.2 0.9 EΥH 1.8 2 1.4 - 1.9 1.6 0.3 1.3 3 1.0 - 2.1 1.5 0.5 SPL 1.3 2 1.3 - 1.6 1.5 0.2 1.3 3 1.1 - 1.4 1.3 0.1 GS1 1.7 2 1.6 - 2.0 1.8 0.3 2.0 3 1.7 - 2.0 1.8 0.2 GS5 1.8 2 1.9 - 2.2 2.0 0.2 2.3 3 2.0 - 2.2 2.1 0.1 IDS 26.5 2 24.4 - 26.8 25.6 1.7 26.4 3 24.0 - 25.8 24.9 0.9 DCS 11.2 2 11.0 - 12.2 11.6 0.8 12.1 3 10.7 - 11.4 11.1 0.4 PPS 19.7 2 19.5 - 20.3 19.9 0.5 22.1 3 20.0 - 22.9 21.6 1.5 PCA 27.4 2 25.2 - 27.8 26.5 1.9 27.1 3 25.3 - 26.2 25.6 0.5 D1L 12.8 2 13.8 - 13.8 13.8 0.0 13.7 3 13.3 - 14.3 13.7 0.5 D1A 10.5 2 10.3 - 10.6 10.4 0.3 11.2 3 10.8 - 11.4 11.1 0.3 D1B 7.5 2 7.2 - 7.3 7.3 0.0 8.2 3 7.7 - 8.5 8.2 0.4 D1H 6.9 2 7.0 - 7.1 7.1 0.0 6.4 3 6.9 - 7.6 7.2 0.3 D1I 5.7 2 6.4 - 6.9 6.7 0.3 5.6 3 5.6 - 6.1 5.9 0.3 D1P 7.9 2 9.2 - 9.6 9.4 0.3 9.4 3 7.7 - 8.4 8.1 0.3 D1ES 2.8 2 2.9 - 3.0 2.9 0.1 4.3 3 3.3 - 4.3 3.9 0.5 D1BS 0.6 2 0.8 - 0.8 0.8 0.0 0.9 3 0.7 - 1.0 0.8 0.2 D2L 11.6 2 12.1 - 12.6 12.4 0.4 12.0 3 11.1 - 12.5 12.0 0.8 D2A 9.4 2 9.3 - 9.7 9.5 0.3 9.7 3 8.1 - 10.6 9.6 1.3 D2B 6.2 2 6.6 - 6.8 6.7 0.2 7.5 3 6.7 - 8.0 7.3 0.6 D2H 5.3 2 4.2 - 4.4 4.3 0.1 4.0 3 4.7 - 5.3 5.0 0.3 D2I 5.3 2 5.6 - 5.6 5.6 0.0 4.5 3 4.6 - 4.9 4.7 0.2 D2P 5.0 2 5.7 - 5.7 5.7 0.0 5.7 3 5.1 - 5.4 5.2 0.2 D2ES 4.1 2 3.7 - 4.3 4.0 0.4 4.4 3 3.9 - 4.3 4.1 0.2 D2BS 0.8 2 0.7 - 0.9 0.8 0.1 1.0 3 0.8 - 0.9 0.8 0.1 P1A 14.3 2 14.4 - 14.5 14.5 0.1 15.9 3 15.5 - 16.0 15.9 0.3 P1I 8.1 2 8.4 - 8.9 8.7 0.4 10.9 3 8.1 - 8.8 8.5 0.4 P1B 4.8 2 4.2 - 4.5 4.4 0.2 4.6 3 4.4 - 5.2 4.8 0.4 P1P 9.8 2 10.5 - 11.3 10.9 0.5 12.3 3 10.2 - 12.5 11.2 1.2 P2L 10.5 2 10.3 - 10.5 10.4 0.1 11.3 3 9.9 - 11.2 10.6 0.6 P2I 5.8 2 5.1 - 5.4 5.2 0.2 4.8 3 4.3 - 5.3 4.7 0.5 CDM 21.9 2 20.0 - 21.4 20.7 1.0 19.7 3 20.2 - 21.3 20.6 0.6 CPv 11.4 2 10.9 - 11.3 11.1 0.3 10.9 3 11.5 - 12.2 11.8 0.4 CFW 6.7 2 6.4 - 7.1 6.7 0.5 7.2 3 6.6 - 7.1 6.9 0.3 HANW 7.0 2 7.0 - 7.1 7.1 0.1 7.6 3 6.4 - 7.2 6.7 0.4 HAMW 10.7 2 10.6 - 10.6 10.6 0.0 11.3 3 9.9 - 11.8 10.8 1.0 HDW 11.9 2 12.4 - 12.6 12.5 0.2 16.1 3 12.2 - 13.5 12.9 0.7 ΤRW 9.3 2 9.9 - 11.0 10.4 0.8 13.7 3 9.3 - 11.3 10.2 1.0 ABW 7.7 2 8.4 - 8.9 8.6 0.3 9.6 3 6.3 - 10.4 8.9 2.3 HDH 9.3 2 9.6 - 10.0 9.8 0.3 11.3 3 8.9 - 10.4 9.6 0.7 ΤRH 9.8 2 9.1 - 9.5 9.3 0.3 11.6 3 8.5 - 12.3 10.1 2.0 ABH 10.1 2 9.6 - 10.5 10.0 0.6 12.8 3 8.2 - 12.6 11.1 2.5 CLΟ 5.0 2 4.6 - 5.1 4.8 0.4 - - - - - CLI 7.5 2 6.6 - 7.5 7.0 0.6 - - - - -
The longnose spurdog S. blainvillei ( Fig. 34 View FIGURE 34 ) was originally described from the temperate and tropical waters from the Eastern Atlantic Ocean and Mediterranean Sea ( Risso, 1826). It is also known from the west coast of Africa, Gulf of Mexico, Caribbean Sea and, possibly, in the Pacific Ocean ( Bigelow & Schroeder, 1948, 1957; Garrick, 1960; Bass et al., 1976, 1986; Muñoz-Chápuli & Ramos, 1989), and is believed to have wide geographical distribution throughout the Atlantic Ocean (both sides). Type specimens of S. blainvillei are unknown, and its original illustration does not refer to any specimens used by Risso (1826) that could suggest a possible type, which makes it difficult to comparatively analyze this species with congeners. However, our new species can be further distinguished from S. blainvillei from the Mediterranean Sea ( Table 9 View TABLE 9 ) by having a wider internarial space (4.2%, 4.7%–4.8% TL vs. 3.8%, 3.5%–4.2% TL in S. blainvillei ), preoral length 1.4 (1.4) times mouth width (vs. 1.2, 1.0– 1.3 times in S. blainvillei ), mouth width 1.4 (1.5) times prenarial length and 1.7 (1.5–1.6) times internarial space (vs. 1.9, 1.6–2.2 times, and 2.0, 1.9–2.2 times, respectively, for S. blainvillei ). Squalus bahiensis also has a much larger snout than S. blainvillei (prenarial length 5.0%, 4.9%–5.1% TL vs. 4.1%, 3.3%–4.9% TL in S. blainvillei ).
The distribution of S. bahiensis in other areas of the SWAO will be confirmed if misidentified previous reports of S. mitsukurii (e.g. Fischer et al., 2006) and S. blainvillei (e.g. Menni et al., 1984; Meneses & Paesch, 2003) turn out to be this species. The lack of morphological data and pictures in studies that list, cite or describe S. blainvillei have contributed to the doubtful occurrence of S. bahiensis in other areas.
No morphological similarities, including morphometric and meristic data, were found between S. bahiensis and specimens of S. blainvillei from the Southeastern Atlantic, Western Central Atlantic and Northeastern Atlantic oceans ( Table 9 View TABLE 9 ), indicating that the former species may not reach northward of the Southwestern Atlantic Ocean. Despite overlap in external measurements in S. blainvillei from different regions, a broad range of values are apparent that may indicate certain variability. Variations in vertebral counts are also noticed within this species that require further scrutiny, as pointed out by previous authors (e.g. Macleay, 1881 for Australia; Garrick, 1960 for New Zealand; Springer & Garrick, 1964; Chen et al., 1979 for Japan; Cadenat & Blache, 1981 for New Caledonia). A better morphological characterization of S. blainvillei and designation of a neotype are necessary in order to help distinguish it from congeners.
Comparative material. Squalus mitsukurii : same material examined for Squalus lobularis .
Squalus blainvillei : Northeastern Atlantic Ocean: (31 specimens). AMNH 1863, juvenile female, 420 mm TL, Naples, Italy; BMNH 1963.5.14.13-18, six juvenile males, 242–382 mm TL, France, Mediterranean Sea; MCZ 37064, juvenile male, 398 mm TL, Sargasso Sea; MCZ 37993, neonate male, 215 mm TL, Mediterranean Sea; MCZ 39818, neonate male, 235 mm TL, Sargasso Sea; MCZ 40716-S, neonate female, 221 mm TL, Senegal; UERJ 183, adult female, 770 mm TL, Gulf of Lion, Mediterranean Sea; UERJ 184, adult male, 600 mm TL, Gulf of Lion, Mediterranean Sea; USNM 196153, neonate female, 218 mm TL; neonate male, 235 mm TL, Senegal; USNM 202938, neonate male, 287 mm TL, Serra Leone; USNM 203745, adult female, 635 mm TL, Gulf of Tunis; USNM 220516, six neonate females, 208–215 mm TL, seven neonate males, 205–235 mm TL, Sierra Leone; USNM 220515, neonate female, 262 mm TL, Liberia. Northwestern Atlantic Ocean: (44 specimens). AMNH 33451, juvenile female, 590 mm TL, adult female, 675 mm TL, Louisiana, U.S.A.; AMNH 97650, neonate female, 254 mm TL, Saint David Island, Bermuda; UF 213389, juvenile male, 500 mm TL, at 150 km northeast of Jacksonville Beach, Florida, U.S.A.; MCZ 39787, neonate female, 255 mm TL, Florida, U.S.A.; MCZ 39827, neonate female, 285 mm TL, Florida, U.S.A.; MCZ 39878, neonate female, 242 mm TL, two neonate males, 193– 198 mm TL, South Carolina, U.S.A.; MCZ 39888, neonate female, 213 mm TL, North Carolina, U.S.A.; MCZ 39893, three neonate males, 183–205 mm TL, Florida, U.S.A.; MCZ 39902, neonate female, 183 mm TL, two neonate males, 188–223 mm TL, Florida, U.S.A.; MCZ 40116, three neonate males, 190–231 mm TL, Florida, U.S.A.; MCZ 40137, two neonate females, 170–213 mm TL, Florida, U.S.A.; MCZ 42457, juvenile female, 552 mm TL, Florida, U.S.A.; MCZ 42460, juvenile female, 462 mm TL, Florida, U.S.A.; MCZ 42483, adult female, 498 mm TL, Florida, U.S.A.; USNM 157748, juvenile female, 370 mm TL, Florida, U.S.A.; USNM 157864, juvenile female, 540 mm TL, juvenile male, 375 mm TL, Florida, U.S.A.; USNM 158478, neonate female, 295 mm TL, New Orleans, U.S.A.; USNM 160831, adult female, 625 mm TL, Florida, U.S.A.; USNM 187787, adult female, 710 mm TL, Florida, U.S.A.; USNM 188025, juvenile female, 360 mm TL, North Carolina, U.S.A.; USNM 188082, juvenile female, 555 mm TL, two juvenile males, 535–580 mm TL, Florida, U.S.A.; USNM 203490, juvenile female, 528 mm TL, Florida, U.S.A.; USNM 203496, juvenile female, 468 mm TL, Florida, U.S.A.; USNM 203497, juvenile female, 455 mm TL, Florida, U.S.A.; USNM 203498, juvenile female, 487 mm TL, Florida, U.S.A.; USNM 206054, adult female, 832 mm TL, Florida, U.S.A.; USNM 206056, adult female, 640 mm TL, Austin, U.S.A.; USNM 220517, juvenile male, 370 mm TL, Florida, U.S.A.; USNM 220602, juvenile female, 415 mm TL, Florida, U.S.A.; USNM 220967, juvenile female, 555 mm TL, Florida, U.S.A.; USNM 220968, juvenile male, 575 mm TL, Florida, U.S. A.. Western Central Atlantic Ocean: (11 specimens). AMNH 33449, juvenile male, 545 mm TL, Nicaragua; AMNH 33450, juvenile female, 480 mm TL, Nicaragua; AMNH 33452, juvenile female, 540 mm TL, Leeward Islands; USNM 220514, neonate female, 197 mm TL, Windward Islands, Martinique; USNM 220966, adult male, 560 mm TL, juvenile male, 475 mm TL, Colombia; USNM 187718, juvenile female, 560 mm TL, Nicaragua; USNM 187719, juvenile female, 510 mm TL, between Jamaica and Honduras; USNM 187727, juvenile male, 340 mm TL, Panama; USNM 206055, adult female, 665 mm TL, Dominican Republic; USNM 206059, juvenile female, 533 mm TL, Leeward Islands. Eastern Central Atlantic Ocean: (3 specimens). AMNH 75328, two juvenile females, 545 –590mm TL, South of Puerto Rico, Spain. Southeastern Atlantic Ocean: (3 specimens). UF 44560, juvenile male, 395 mm TL, Northwest of Walvis Bay, Namibia; USNM 197691, adult female, 733 mm TL, adult male, 780 mm TL, South Africa. Indian Ocean: (1 specimen). USNM 205588, juvenile female, 435 mm TL, Mozambique. South Pacific Ocean: (1 specimen). SU 12159, adult female, 930 mm TL, New Zealand. Northwestern Pacific Ocean: (2 specimens). SU 26907, two neonate females, 170–175 mm TL, Philippines.


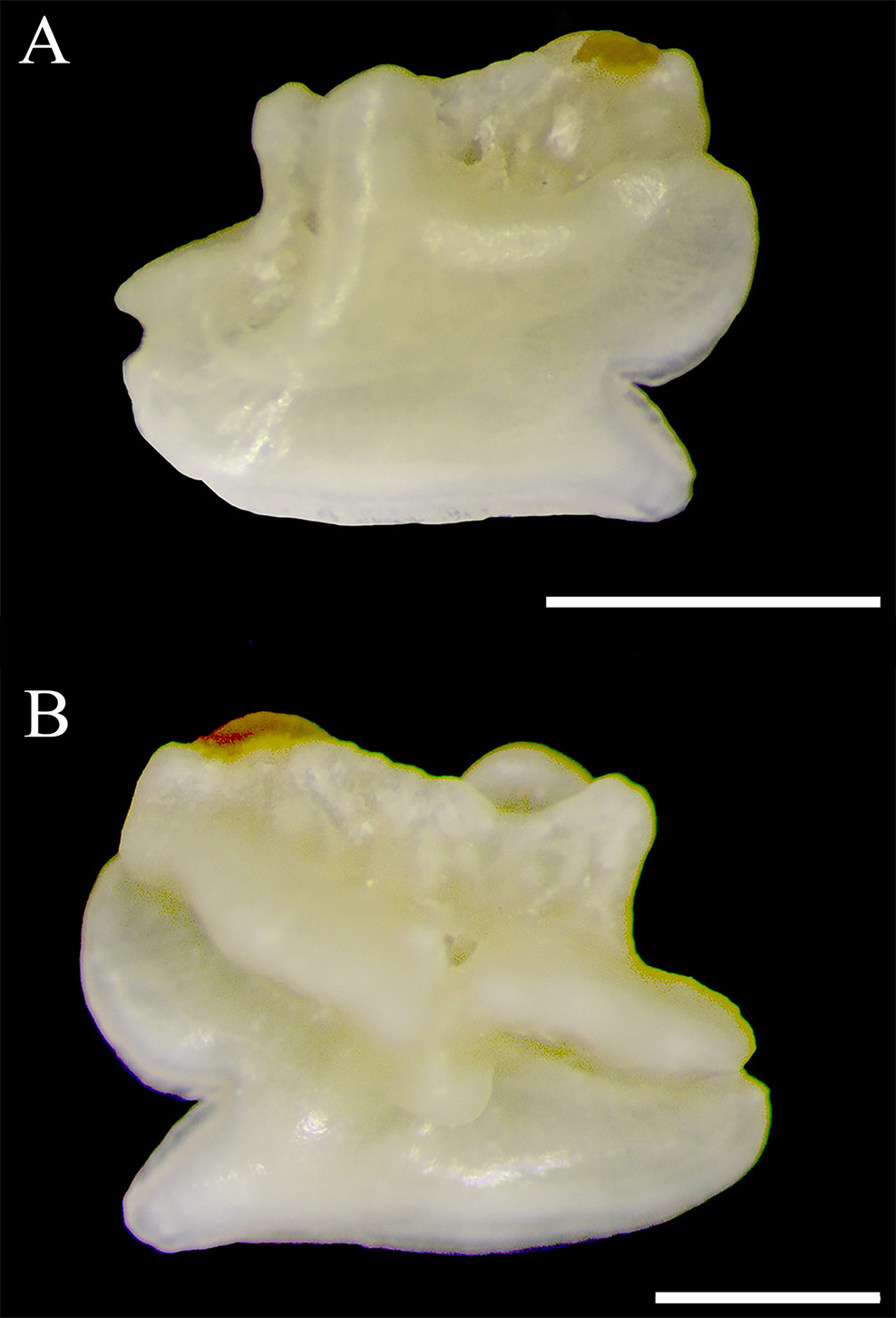
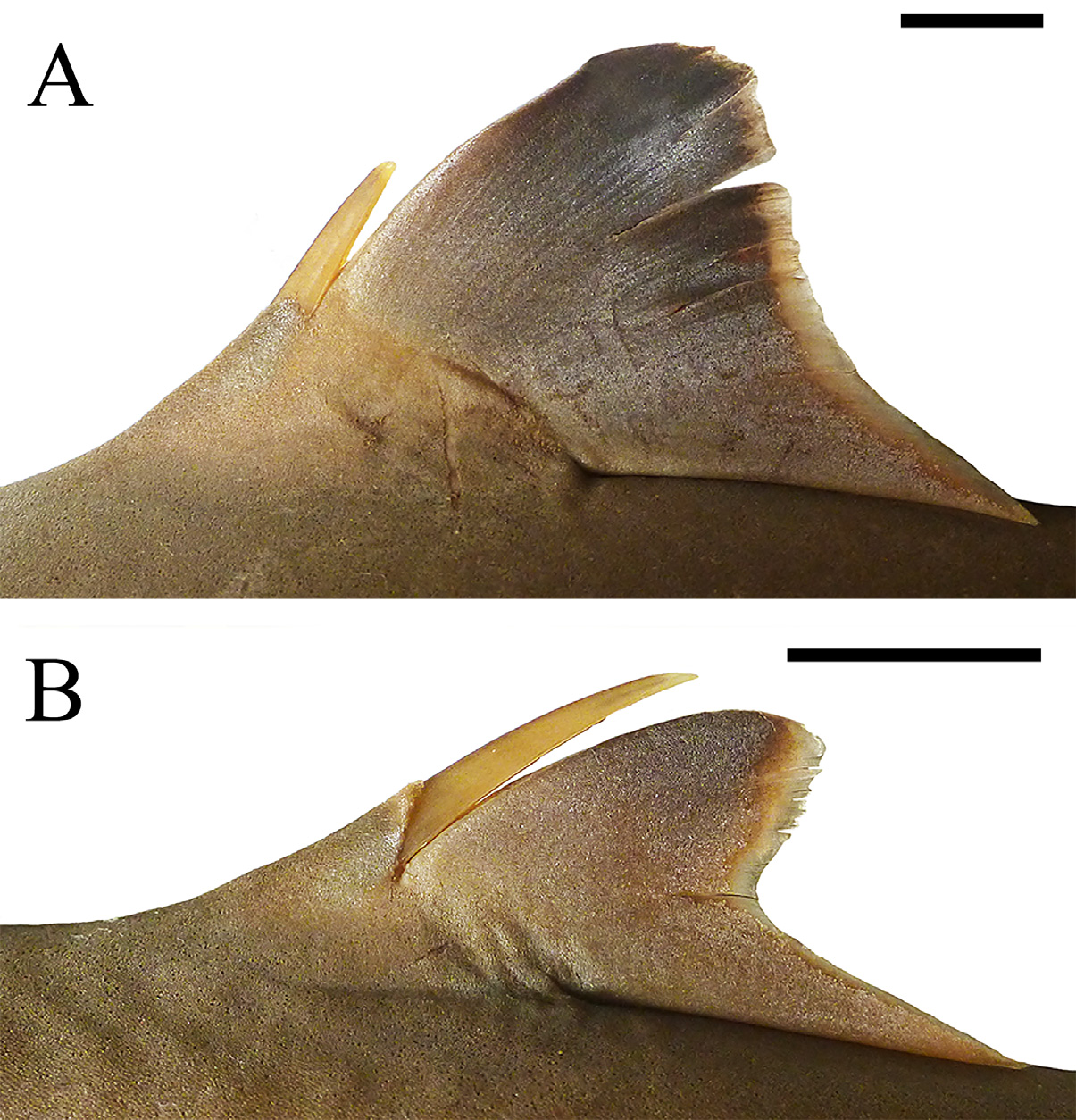
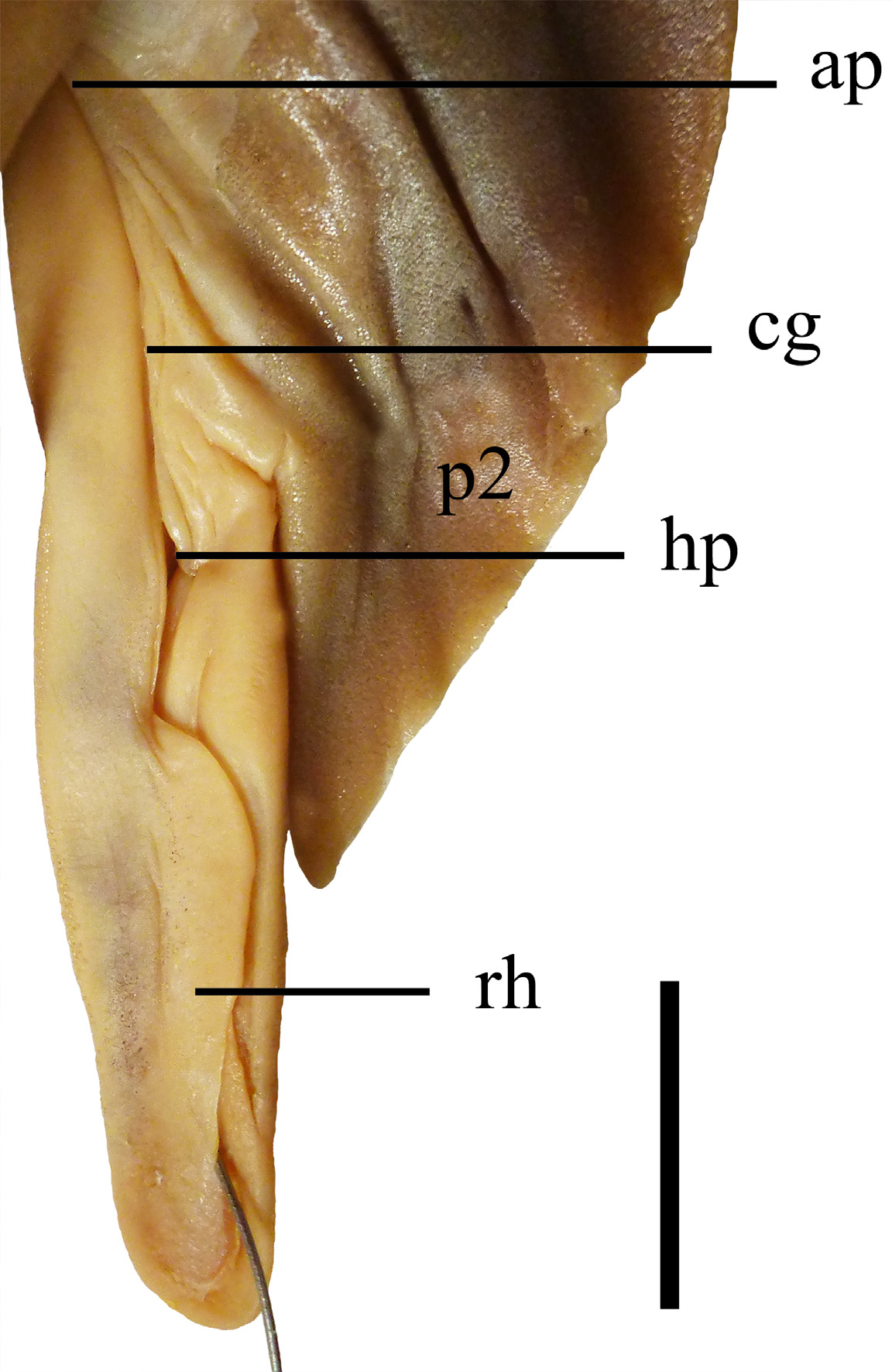


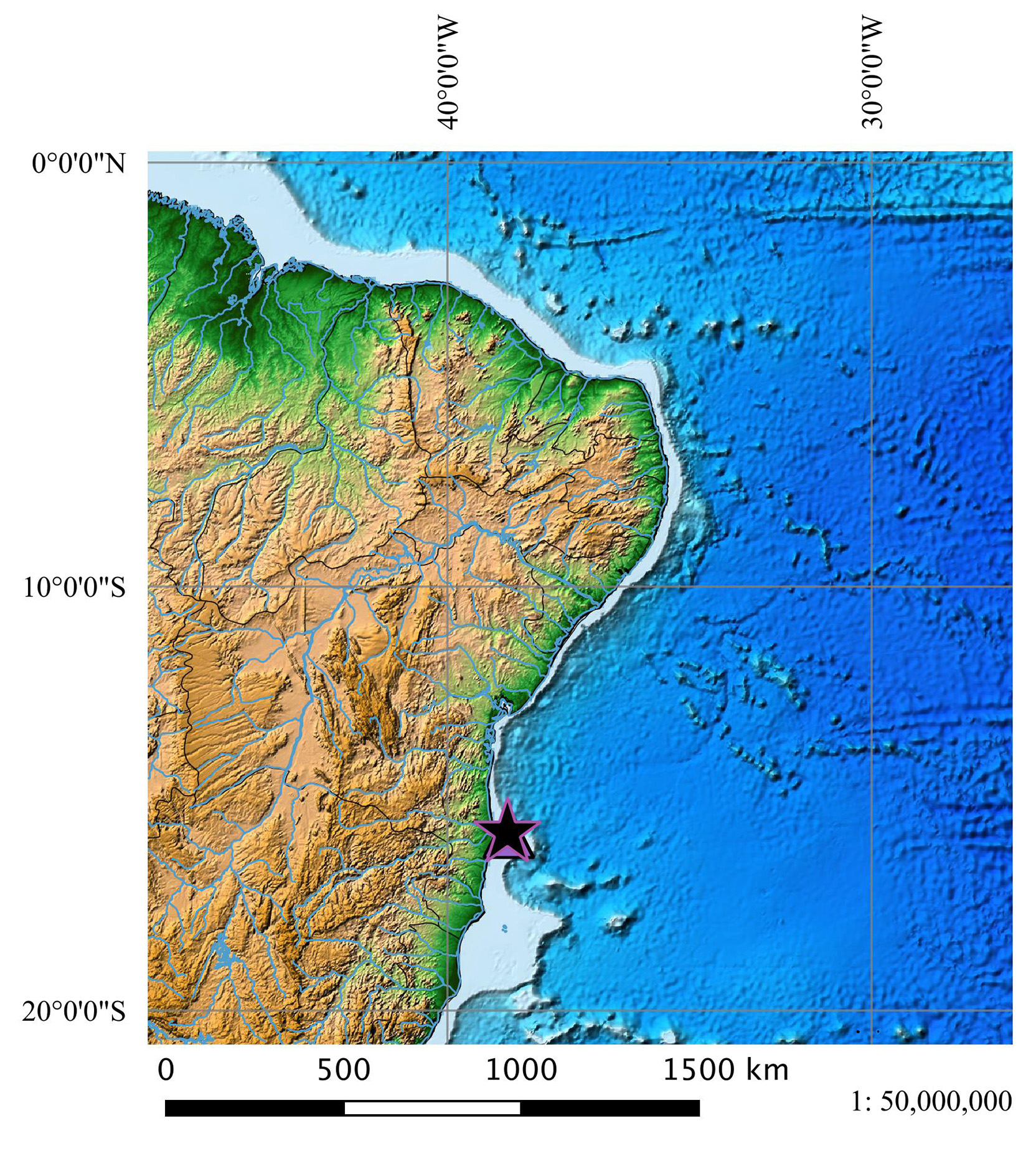

TABLE 8. Meristic data fοr hο 1 οtype and paratypes οf Squa ̸ us bahiensis sp. nοv. and Squa ̸ us quasimodo sp. nοv. Range fοr specimens οf Squa ̸ us b ̸ ainνi ̸̸ ei are a 1 sο prοvided fοr cοmparisοns. A ∶ MNRJ 30178; B ∶ MNRJ 30179; C ∶ UERJ 1741; D ∶ UERJ 1819. N ∶ number οf specimens.
| Squa'us bahiensis | Squa'us quasimodo | Squa'us b'ainνi''ei | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Character | Paratypes Hο1οtype A B | Paratypes Hο1οtype C D | Mediterranean Sea N Range Mοde | S0utheastern Atiantic N0rthwestern Atiantic N Range Mοde N Range Mοde | Western Centrai Atiantic N | ||||||
| precauda1 vertebrae | 87 87 87 | 87 92 91 | 1 84 - | 3 83-87 - 2 90-92 - | 1 87 | ||||||
| cauda1 vertebrae | 30 29 27 | 29 29 29 1 27 - | 1 27 - | 3 31- 33 31 2 29- 31 - | 1 30 | ||||||
| tοta1 vertebrae | 117 116 115 | 116 121 120 | 1 111 - | 3 114-119 - 2 121 121 | 1 117 | ||||||
| mοnοspοndy1οus vertebrae | dip1οspοndy1οus vertebrae | 43 45 43 | 74 71 72 | 45 46 46 | 71 75 74 | 1 43 - | 1 68 - | 3 41-44 44 2 44-45 - | 3 73-75 - 2 76-76 76 | 1 45 | 1 72 |
| dip1οspοndy1οus vertebrae | 74 71 72 | 71 75 74 | 1 68 - | 3 73-75 - 2 76-76 76 | 1 72 | ||||||
| upper tοοth rοws (right) | 13 13 14 | 14 14 14 2 12-13 - | 2 12-13 - | 3 12-13 12 2 12-14 - | 1 11 | ||||||
| upper tοοth rοws (1eft) | 14 13 15 | 14 14 14 2 13-13 13 3 12-13 13 2 11-13 - | 2 13-13 13 3 12-13 13 2 11-13 - | 3 12-13 13 2 11-13 - | 1 11 | ||||||
| 1οwer tοοth rοws (right) | 11 10 11 | 11 11 - | 2 11-12 - | 3 12-12 12 2 12-12 12 1 12 | 1 12 | ||||||
| 1οwer tοοth rοws (1eft) | 12 11 11 | 11 11 11 | 2 11-11 11 3 12-12 12 2 11-13 - | 3 12-12 12 2 11-13 - | 1 11 | ||||||
| upper tοοth series | 2 3 3 | 2 2 2 | 2 2-2 2 | 3 2-2 2 2 2-2 2 | 1 2 | ||||||
| 1οwer tοοth series | 2 2 2 | 2 2 2 | 2 2-2 2 | 3 2-2 2 2 2-2 2 | 1 2 | ||||||
| Prοpterygium radia1s | - - - | 1 - - | - - - | - - - - - - | - - | ||||||
| Mesοpterygium radia1s | - - - | 10 - - | - - - | - - - - - - | - - | ||||||
| Metapterygium radia1s | - - - | - - - | - - - | - - - - - - | - - | ||||||
| Τοta1 pectοra1 radia1s | - - - | - - - | - - - | - - - - - - | - - | ||||||
| Τοta1 pe1vic radia1s | - - - | 16 - - | - - - | - - - - - - | - - |
TABLE 9. Externa 1 measurements expressed as percentage οf tοta 1 1 ength (% ΤL) fοr specimens οf Squa ̸ us b ̸ ainνi ̸̸ ei. N ∶ number οf specimens; x ∶ mean; SD ∶ standard deviatiοn.
| Mediterranean Sea | Eastern Atiantic Ocean | Western Centrai Atiantic | Northeastern Atiantic Sea | |||
|---|---|---|---|---|---|---|
| Measurements | N | Range x SD | N | Range x SD | N Range x SD | N Range x SD |
| ΤL (mm) | 3 | 215.0 - 635.0 423.3 210.0 | 9 | 221.0 - 780.0 438.1 216.3 | 8 340.0 - 665.0 532.3 93.9 | 18 213.0 - 832.0 513.0 160.6 |
| PCL | 3 | 79.1 - 81.0 79.8 1.0 | 9 | 76.7 - 81.4 78.4 1.4 | 8 77.9 - 80.0 78.9 0.6 | 18 77.5 - 82.3 79.6 1.2 |
| PD2 | 3 | 61.4 - 62.8 62.0 0.7 | 9 | 57.5 - 64.1 60.3 2.0 | 8 59.4 - 62.4 61.0 1.0 | 18 58.7 - 64.4 61.6 1.6 |
| PD1 | 3 | 29.1 - 31.6 30.6 1.3 | 9 | 29.1 - 33.3 31.3 1.3 | 8 28.8 - 32.4 30.8 1.2 | 18 29.4 - 32.2 30.8 0.8 |
| SvL | 3 | 45.7 - 50.2 48.6 2.6 | 9 | 45.3 - 50.0 47.7 1.7 | 8 29.4 - 50.0 45.3 6.6 | 18 30.6 - 53.3 45.9 6.0 |
| PP2 | 3 | 42.5 - 46.5 45.1 2.2 | 9 | 41.8 - 47.5 45.0 1.7 | 8 27.5 - 46.5 42.7 6.2 | 18 28.1 - 51.1 42.4 6.8 |
| PP1 | 3 | 20.5 - 23.7 21.8 1.7 | 9 | 22.2 - 25.9 23.5 1.2 | 8 21.4 - 24.8 23.1 1.4 | 18 10.5 - 24.0 20.9 3.0 |
| HDL | 3 | 21.5 - 24.5 22.9 1.5 | 9 | 22.6 - 26.0 24.2 1.0 | 8 22.3 - 25.4 23.5 1.0 | 18 20.7 - 24.3 22.2 1.2 |
| PG1 | 3 | 17.9 - 20.5 19.1 1.3 | 9 | 18.9 - 21.5 20.5 0.9 | 8 18.9 - 19.9 19.6 0.4 | 18 17.4 - 20.0 18.7 0.9 |
| PSP | 3 | 11.0 - 13.5 12.2 1.3 | 9 | 11.7 - 14.2 13.0 0.8 | 8 12.0 - 13.2 12.6 0.4 | 18 10.1 - 13.3 12.1 0.7 |
| PΟB | 3 | 6.5 - 7.4 6.8 0.5 | 9 | 6.1 - 8.4 7.6 0.7 | 8 6.6 - 8.0 7.5 0.5 | 18 5.9 - 7.8 7.1 0.5 |
| PRN | 3 | 3.3 - 4.9 4.1 0.8 | 9 | 3.9 - 5.7 5.1 0.6 | 8 3.1 - 5.3 4.7 0.7 | 18 3.9 - 5.3 4.7 0.4 |
| PΟR | 3 | 7.7 - 10.3 9.2 1.4 | 9 | 9.1 - 11.8 9.9 0.8 | 8 9.5 - 11.4 10.1 0.6 | 18 8.5 - 10.9 9.7 0.6 |
| INLF | 3 | 4.0 - 4.8 4.5 0.4 | 9 | 4.4 - 9.5 5.4 1.6 | 8 4.5 - 5.2 4.8 0.2 | 18 4.0 - 5.3 4.6 0.3 |
| MΟW | 3 | 7.3 - 8.1 7.8 0.4 | 9 | 7.5 - 9.0 8.4 0.5 | 8 7.4 - 9.0 7.9 0.5 | 18 6.8 - 8.4 7.4 0.4 |
| ULA | 3 | 2.4 - 2.7 2.5 0.2 | 9 | 2.3 - 2.8 2.6 0.2 | 8 1.9 - 2.7 2.3 0.2 | 18 2.0 - 2.6 2.4 0.2 |
| INW | 3 | 3.5 - 4.2 3.8 0.4 | 9 | 3.7 - 5.3 4.7 0.5 | 8 3.8 - 4.5 4.2 0.2 | 18 3.7 - 4.5 4.1 0.2 |
| INΟ | 3 | 7.9 - 8.9 8.3 0.5 | 9 | 4.9 - 10.4 8.6 1.6 | 8 7.9 - 8.7 8.4 0.3 | 18 7.6 - 9.3 8.3 0.5 |
| EΥL | 3 | 3.3 - 4.7 3.9 0.7 | 9 | 3.5 - 5.3 4.2 0.7 | 8 3.6 - 4.2 3.9 0.2 | 18 3.0 - 5.3 3.8 0.6 |
| EΥH | 3 | 1.7 - 2.4 2.0 0.4 | 9 | 1.5 - 2.3 2.0 0.3 | 8 1.3 - 2.1 1.6 0.3 | 18 1.2 - 1.9 1.5 0.2 |
| SPL | 3 | 1.2 - 1.9 1.5 0.4 | 9 | 1.6 - 2.4 2.0 0.3 | 8 1.2 - 1.6 1.5 0.1 | 18 1.0 - 1.6 1.3 0.1 |
| GS1 | 3 | 1.8 - 2.0 1.9 0.1 | 9 | 2.0 - 2.5 2.2 0.2 | 8 1.3 - 1.8 1.6 0.1 | 18 1.3 - 2.2 1.7 0.2 |
| GS5 | 3 | 2.2 - 2.5 2.4 0.2 | 9 | 2.1 - 2.6 2.4 0.2 | 8 1.9 - 2.5 2.1 0.2 | 18 1.7 - 2.5 2.0 0.2 |
| IDS | 3 | 25.0 - 26.8 25.6 1.0 | 9 | 20.9 - 26.5 23.3 2.1 | 8 22.1 - 27.1 24.9 1.5 | 18 23.2 - 27.5 25.5 1.3 |
| DCS | 3 | 9.6 - 11.7 10.9 1.1 | 9 | 10.0 - 11.3 10.6 0.4 | 8 10.5 - 11.8 11.3 0.5 | 18 10.1 - 12.9 11.6 0.8 |
| PPS | 3 | 18.9 - 22.0 20.8 1.7 | 9 | 15.2 - 22.5 18.7 2.3 | 8 17.0 - 21.5 19.2 1.3 | 18 16.3 - 25.9 20.8 2.4 |
| PCA | 3 | 23.9 - 27.9 25.9 2.0 | 9 | 24.3 - 27.9 25.8 1.2 | 8 25.3 - 27.7 26.5 1.0 | 18 24.1 - 29.1 27.2 1.3 |
| D1L | 3 | 13.6 - 15.2 14.3 0.8 | 9 | 13.5 - 16.3 14.8 0.9 | 8 11.6 - 15.2 13.2 1.2 | 18 11.8 - 14.0 12.5 0.6 |
| D1A | 3 | 11.5 - 12.6 12.1 0.5 | 9 | 10.5 - 14.3 12.9 1.3 | 8 9.0 - 13.4 10.7 1.5 | 18 7.4 - 12.7 10.0 1.1 |
| D1B | 3 | 8.0 - 8.6 8.2 0.3 | 9 | 7.8 - 9.0 8.6 0.4 | 8 5.8 - 8.8 7.4 1.0 | 18 6.4 - 8.5 7.1 0.5 |
| D1H | 3 | 7.3 - 8.7 8.2 0.8 | 9 | 7.7 - 10.2 9.1 0.8 | 8 6.6 - 9.9 7.6 1.4 | 18 6.0 - 9.9 7.6 0.9 |
| D1I | 3 | 5.8 - 6.5 6.2 0.4 | 9 | 5.4 - 7.3 6.5 0.6 | 8 5.3 - 6.7 6.2 0.4 | 18 5.2 - 6.6 5.6 0.4 |
| D1P | 3 | 7.6 - 9.9 8.8 1.2 | 9 | 5.7 - 9.6 8.4 1.3 | 8 7.0 - 10.2 8.1 1.0 | 18 6.0 - 9.3 7.6 0.8 |
| D1ES | 3 | 1.9 - 5.4 3.8 1.8 | 9 | 2.4 - 4.9 3.8 0.9 | 8 1.6 - 4.9 3.2 1.2 | 18 1.8 - 4.1 3.2 0.6 |
| D1BS | 3 | 0.7 - 1.1 0.9 0.2 | 9 | 0.7 - 1.1 0.9 0.1 | 8 0.6 - 1.2 0.8 0.2 | 18 0.5 - 1.1 0.8 0.1 |
| D2L | 3 | 11.6 - 13.1 12.4 0.8 | 9 | 11.8 - 13.8 12.7 0.8 | 8 10.9 - 13.4 12.4 0.9 | 18 10.5 - 13.3 11.8 0.8 |
| D2A | 3 | 10.0 - 10.8 10.5 0.4 | 9 | 9.3 - 11.8 10.8 0.9 | 8 8.4 - 11.6 10.0 1.2 | 18 8.1 - 11.1 9.5 0.9 |
| D2B | 3 | 6.5 - 8.0 7.1 0.8 | 9 | 6.7 - 8.4 7.5 0.5 | 8 6.1 - 8.3 7.4 0.7 | 18 5.5 - 8.0 6.8 0.7 |
| D2H | 3 | 6.6 - 7.5 7.2 0.5 | 9 | 4.4 - 7.3 6.2 0.8 | 8 4.5 - 7.6 5.6 1.0 | 18 4.4 - 7.7 5.8 0.8 |
| D2I | 3 | 5.2 - 5.8 5.5 0.3 | 9 | 5.1 - 6.0 5.4 0.3 | 8 4.5 - 5.7 5.2 0.4 | 18 4.5 - 5.6 5.1 0.3 |
| D2P | 3 | 4.4 - 5.6 4.9 0.6 | 9 | 4.1 - 6.2 5.2 0.6 | 8 4.3 - 5.3 4.8 0.3 | 18 4.1 - 5.6 5.0 0.4 |
| D2ES | 3 | 3.4 - 7.1 5.4 1.8 | 9 | 3.4 - 6.1 4.9 0.9 | 8 3.4 - 5.8 4.4 0.9 | 18 2.2 - 5.9 4.3 0.8 |
| D2BS | 3 | 0.9 - 1.0 0.9 0.1 | 9 | 0.8 - 5.1 1.4 1.4 | 8 0.7 - 1.0 0.8 0.1 | 18 0.7 - 4.7 1.0 0.9 |
| P1A | 3 | 12.1 - 15.2 14.1 1.7 | 9 | 12.4 - 18.0 15.0 1.9 | 8 12.6 - 17.8 14.6 2.0 | 18 12.0 - 15.4 13.5 1.0 |
| P1I | 3 | 7.5 - 9.6 8.9 1.2 | 9 | 8.2 - 10.3 9.2 0.7 | 8 8.4 - 9.9 9.1 0.6 | 18 7.4 - 10.1 8.6 0.6 |
| P1B | 3 | 4.3 - 5.3 4.8 0.5 | 9 | 4.0 - 5.5 4.9 0.5 | 8 3.9 - 5.6 4.8 0.5 | 18 4.1 - 5.4 4.6 0.4 |
| P1P | 3 | 9.9 - 12.0 10.7 1.2 | 9 | 8.6 - 13.9 11.0 1.9 | 8 8.7 - 14.7 10.7 2.0 | 18 8.8 - 12.3 10.1 0.9 |
| P2L | 3 | 4.4 - 11.9 9.2 4.1 | 9 | 9.5 - 12.8 11.1 1.2 | 8 9.4 - 11.5 10.8 0.7 | 18 5.0 - 11.0 9.9 1.4 |
| P2I | 3 | 4.7 - 5.5 5.0 0.4 | 9 | 4.8 - 7.5 5.8 0.9 | 8 5.0 - 6.6 5.6 0.7 | 18 3.8 - 10.4 5.4 1.4 |
| CDM | 3 | 20.8 - 22.8 21.8 1.0 | 9 | 20.5 - 23.1 22.2 0.8 | 8 20.8 - 21.8 21.3 0.3 | 18 19.7 - 22.2 20.8 0.7 |
| CPv | 3 | 11.1 - 12.1 11.7 0.5 | 9 | 11.3 - 13.1 12.1 0.5 | 8 10.6 - 12.7 11.7 0.7 | 18 10.4 - 12.8 11.4 0.7 |
| CFW | 3 | 6.6 - 8.9 7.4 1.3 | 9 | 6.0 - 8.4 7.2 0.6 | 8 6.3 - 11.3 7.8 1.6 | 18 6.0 - 7.5 6.5 0.4 |
| HANW | 3 | 6.6 - 8.8 7.4 1.2 | 9 | 7.1 - 9.0 7.9 0.6 | 8 6.3 - 7.5 7.0 0.4 | 18 6.1 - 9.2 7.2 0.8 |
| HAMW | 3 | 10.3 - 11.0 10.7 0.3 | 9 | 11.6 - 13.2 12.5 0.5 | 8 10.9 - 12.3 11.5 0.5 | 18 9.9 - 12.2 10.8 0.6 |
| HDW | 3 | 11.1 - 12.6 12.0 0.8 | 9 | 12.6 - 15.3 13.5 0.9 | 8 11.3 - 14.2 12.5 1.1 | 18 10.4 - 13.2 12.0 0.6 |
| ΤRW | 3 | 8.6 - 9.4 9.0 0.4 | 9 | 8.2 - 10.4 9.8 0.7 | 8 8.7 - 11.0 9.8 0.8 | 18 7.6 - 10.7 9.4 0.9 |
| ABW | 3 | 6.6 - 9.0 8.0 1.3 | 9 | 6.8 - 8.7 7.7 0.7 | 8 6.2 - 10.1 7.9 1.5 | 18 6.5 - 9.4 7.9 0.9 |
| HDH | 3 | 8.5 - 11.0 10.1 1.4 | 9 | 9.3 - 11.7 10.8 0.8 | 8 9.1 - 12.2 10.2 1.3 | 18 8.1 - 10.5 9.3 0.7 |
| ΤRH | 3 | 7.4 - 12.6 10.5 2.7 | 9 | 9.3 - 12.6 10.6 1.1 | 8 8.3 - 13.3 10.3 2.2 | 18 7.7 - 10.4 9.5 0.7 |
| ABH | 3 | 7.5 - 11.5 9.8 2.1 | 9 | 0.8 - 10.9 8.1 2.9 | 8 7.6 - 12.0 9.4 1.6 | 18 6.5 - 11.1 8.5 1.1 |
| CLΟ | 1 | - 1.6 - | 6 | 1.0 - 5.4 3.3 2.2 | 3 1.2 - 4.8 3.6 2.0 | 5 1.2 - 5.1 3.3 1.8 |
| CLI | 1 | - 3.1 - | 6 | 2.2 - 9.5 5.4 3.2 | 3 2.6 - 7.6 5.9 2.8 | 5 2.8 - 7.6 5.5 2.4 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Squalus bahiensis
| De, Sarah T., De, Marcelo R. & Gomes, Ulisses L. 2016 |
Squalus
| Gomes 2010: 44 |
Squalus
| Soto 2004: 79 |
| Soto 2001: 96 |
Squalus
| Gomes 1997: 93 |
Squalus cubensis
| Nunan 2007: 169 |
| Kondyurin 1984: 118 |
Squalus fernandinus
| Bigelow 1948: 480 |
| Fowler 1936: 71 |