
Potamonautes mariepskoppie, Daniels & Barnes & Marais & Gouws, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.782.1591 |
|
publication LSID |
lsid:zoobank.org:pub:EDDB2243-2EEB-4297-86CE-116AADA37B12 |
|
DOI |
https://doi.org/10.5281/zenodo.5770280 |
|
persistent identifier |
https://treatment.plazi.org/id/C0AE6C3E-5460-450C-87D4-87D24DC47862 |
|
taxon LSID |
lsid:zoobank.org:act:C0AE6C3E-5460-450C-87D4-87D24DC47862 |
|
treatment provided by |
Felipe |
|
scientific name |
Potamonautes mariepskoppie |
| status |
sp. nov. |
Potamonautes mariepskoppie View in CoL sp. nov.
urn:lsid:zoobank.org:act:C0AE6C3E-5460-450C-87D4-87D24DC47862
Figs 1–6 View Fig View Fig View Fig View Fig View Fig View Fig
Diagnosis
Measurements for the holotype as follows: CL = 19.07mm; CWW = 26.76mm; CWP = 11.56 mm; FW = 11.58 mm; PFCD = 2.89 mm; CH = 10.63mm; MCPL = 16.44 mm; MCDL = 10.14 mm; P2ML = 11.06 mm; P2MW = 4.65 mm; P5ML = 9.34 mm; P5MW = 3.61 mm.
CARAPACE. Highly arched ( CH /CL = 0.55) postfrontal crest well-defined, complete, lateral ends meeting anterolateral margins; epigastric crests faint, median sulcus between crests short, not forked posteriorly; exorbital, epibranchial teeth reduced to granules; anterolateral carapace margin with no tooth on epibranchial ( Fig. 2A–C View Fig ).
THIRD MAXILLIPED. Ischium with distinct vertical sulcus; s3/s4 complete, V-shaped, deep, midpoint almost meeting anterior margin of sterno-pleonal cavity; margins of s4 low, not raised ( Fig. 2B View Fig ).
CHELIPED. Dactylus (moveable finger) slim, highly arched, enclosing oval interspace, with three larger teeth interspersed by smaller teeth along length; propodus (fixed finger) with four larger teeth interspersed by smaller teeth along length ( Fig. 2C View Fig ); carpus inner margin distal tooth large, pointed, proximal tooth reduced to granules; medial inferior margin of merus lined with series of small granules terminating distally at small, low distal meral tooth, lateral inferior margin smooth.
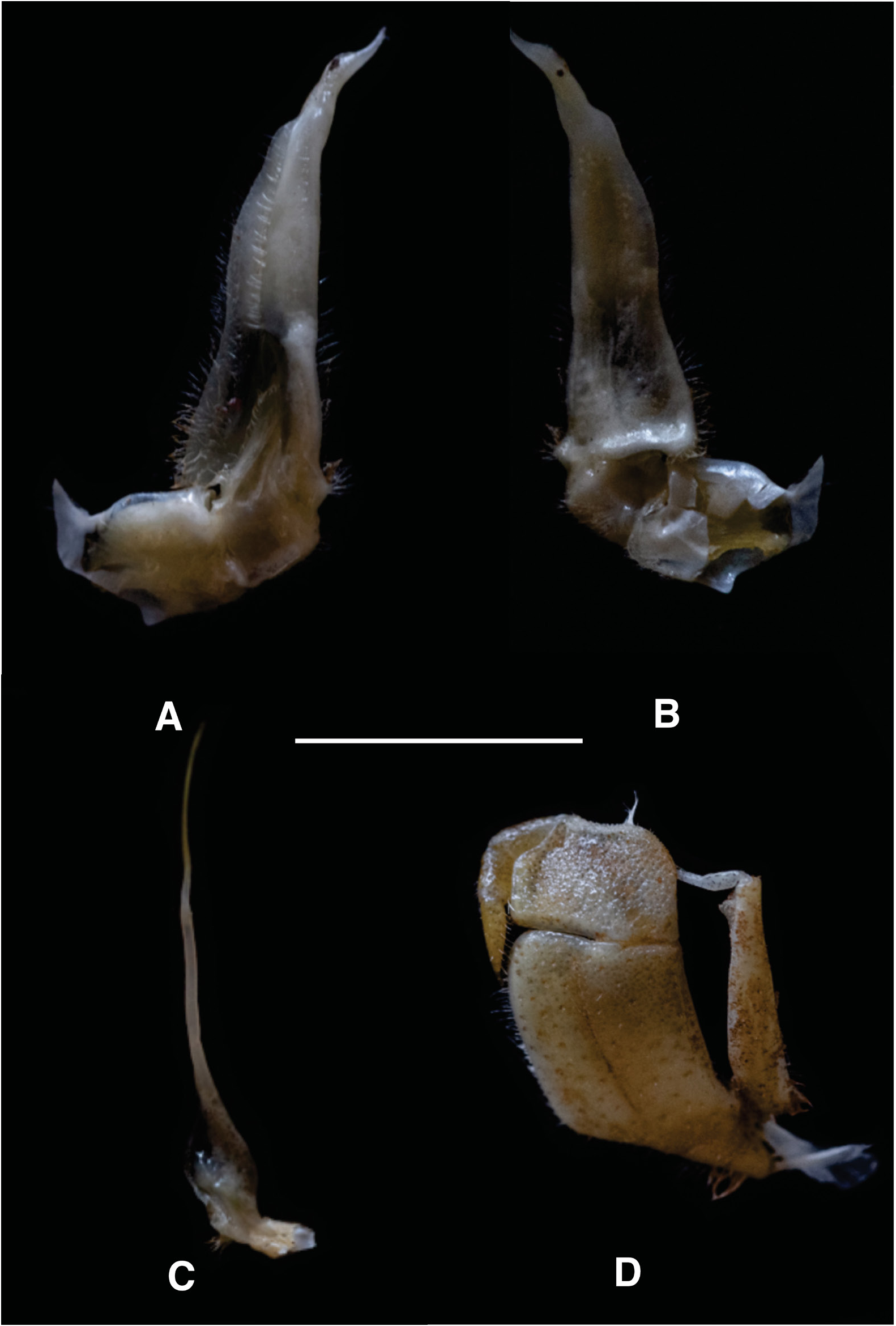
G1 TERMINAL ARTICLE. 1/3 rd length of subterminal; first third straight in line with longitudinal axis of subterminal, middle part directed outward at 45°, widened by raised rounded ventral lobe, tip curving sharply upward ( Fig. 3A–B View Fig ).
Etymology
Named after Mariepskop since it occurs in close proximity to the mountain. The specific epithet is used as a Latin noun in apposition.
Material examined
Holotype SOUTH AFRICA – Mpumalanga Province • ♂; Blyde Canyon Nature Reserve , lowveld section , forest wetland ; 24º42′36.36″ S, 30º54′11.664″ E; 733 m a.s.l.; 21 May 2021; H. Marais leg.; SAM A-094471 . GoogleMaps
Paratype SOUTH AFRICA – Mpumalanga Province • 1 ♂; same collection data as for holotype except 17 Jun. 2020; SAM A-094472 GoogleMaps .
Other material
SOUTH AFRICA – Mpumalanga Province • 4 juvs; same collection data as for paratype; SAM A-094473 GoogleMaps .
Description
Based on the adult male holotype (CWW 26.76 mm).
CARAPACE. Smooth with no tooth on the anteriolateral margins; widest anteriorly, narrowest posteriorly (CWP/CL 0.60); vaulted ( CH /CL 0.55) ( Fig. 2A View Fig ); front broad, one-third of CWW (FW/CWW 0.43); urogastric, cardiac grooves distinct, other grooves faint or missing; postfrontal crest complete, anterolateral margin posterior to epibranchial corner finely granulated, epigastric crests faint, median sulcus between crests short, forked posteriorly; exorbital, epibranchial teeth each reduced to granules; anterolateral margin between exorbital, epibranchial teeth faintly granulated, curving slightly outward, lacking intermediate tooth ( Fig. 2B–C View Fig ); branchiostegal wall vertical sulcus faint, meeting longitudinal sulcus, dividing branchiostegal wall into 3 parts, suborbital, dorsal pterygostomial regions granulated, hepatic region smooth; suborbital margin faintly granulated.
THIRD MAXILLIPED. Filling entire buccal frame, except for respiratory openings; exopod with long flagellum, ischium with faint vertical groove ( Fig. 3D View Fig ). Epistomial tooth large, triangular, margins lined by large granules.
MANDIBLE. Palp two-segmented; terminal simple; tuft of setae at junction between segments.
STERNUM. s1, s2 fused; s2/s3 deep, completely crossing sternum; s3/s4 complete, V-shaped, deep, midpoint almost meeting anterior margin of sterno-pleonal cavity; margins of s4 low, not raised.
CHELIPED. Dactylus (moveable finger) slim, arched, with two teeth interspersed by smaller teeth along length; propodus (fixed finger) with two teeth interspersed by several smaller teeth along length ( Fig. 4A–B View Fig ), tips of both propodus and dactylus black; carpus distal tooth large, pointed, proximal tooth small but distinct, followed by granule; both inferior margins of merus lined with series of small granules, distal meral tooth small, pointed.
PEREOPODS. Walking legs slender, 3 longest, 5 shortest; dorsal margins of pereopods with fine sharp bristles, dactyli of walking legs ending in sharp point, with rows of spine-like bristles.
PLEON. Outline broadly triangular with straight margins. G1 TERMINAL ARTICLE. Short (1/3 length of subterminal), curving away from midline, first third straight in line with longitudinal axis of subterminal segment, middle part directed outward at 45°, widened by low raised rounded ventral lobe, tip curving gently upward. G1 subterminal broad at base, tapering to slim junction with terminal article distally where these two parts have same width, ventral side with heavily setose margins; with setae-fringed flap covering lateral half of segment; dorsal side smooth, no flap, with broad membrane on dorsal side of suture marking junction between terminal, subterminal parts ( Fig. 3A–B View Fig ).
G2 TERMINAL ARTICLE. Long, flagellum-like, 0.5 times length of subterminal ( Fig. 3C View Fig ).
Size
A small-bodied species, CL = 19.07 mm, and wide, CWW = 26.76 mm.
Colour in life
Dorsal carapace chocolate brown with a glossy shine, while the chelipeds and ventral surface are light orange in colour ( Fig. 5A View Fig ).
Distribution
Known only from the lowveld section of the Blyde Canyon Nature Reserve, east of Mariepskop, Mpumalanga Province, South Africa. However, a digital photographic record from iNaturalist (posted by Werner Conradie) suggests the species is also present around Haenertsburg, in Limpopo Province, South Africa ( Fig. 5B View Fig ). Surveys of the latter area are required to confirm this observation.
Ecology
The type locality is located 12 km southeast of Mariepskop Mountain and forms part of the Great Drakensberg Mountain escarpment. The swamp forest receives annual rainfall averaging 1500 mm on top of the mountain and 750mm at the bottom ( Ngwenya et al. 2019). The swamp forests in these areas are all channelled valley-bottom wetlands with connected seeps. The wetland of the type locality is on a shallow gradient that allows the water to seep out slowly into shallow channels and forms shallow muddy puddles, which the crab species prefers. Potamonautes mariepskoppie sp. nov., occurs sympatrically with P. sidneyi sensu stricto. The soils are predominantly organic soils of 15-100cm deep ( Van Rooyen et al. 2020). The presence of large water berries, Syzygium cordatum , is diagnostic of these swamp forests. There are other wetland species present, such as Carex spicata-paniculata , Commelina benghalensis , Cyperus denudatus , Cyclosorus interruptus , Cyperus dives , Isolepis fluitans , Kyllinga odorata , Leersia hexandra , Persicaria decipiens , Selaginella kraussiana , Schoenoplectus brachyceras , Setaria megaphylla , Scleria transvaalensis and Thelypteris confluens .
Remarks
Potamonautes mariepskoppie sp. nov., can be distinguished from its two sister species, P. ngoyensis and P. ntendekaensis , using colour when alive. Potamonautes mariepskoppie sp. nov. has a chocolate brown carapace and orange chelipeds that vary to rust brown as does the specimen from Haenertsburg, Limpopo ( Fig. 5B View Fig ). Potamonautes ngoyensis has a pale-white carapace and chelipeds; P. Potamonautes ntendekaensis has a chocolate-coloured carapace with red pereopods; the entire animal fades to bright orange/red when preserved in absolute ethanol ( Daniels et al. 2019). In addition, P. mariepskoppie sp. nov. is a small-bodied species (CWW = 26.76mm) with a highly arched carapace ( CH /CL = 0.60), while P. ngoyensis is a large-bodied (CWW = 32.8 mm) and flat species ( CH /CL = 0.50). Similarly, P. ntendekaensis is a large-bodied species (CWW = 37.56 mm), that is highly arched ( CH /CL = 0.55). All three species appear to be narrow-endemic forest-dwelling species and are poorly collected based on current distribution records. Potamonautes mariepskoppie sp. nov. appears to be confined to swamp forest patches east of Mariepskop and possibly at Haenertsburg, Limpopo province, while P. ntendekaensis is endemic to the Ntendeka Wilderness (Ngome forest – representing Eastern Scarp forest) area of KwaZulu-Natal, and P. ngoyensis is endemic to the Ngoye forest, part of the greater IOCB forest of KwaZulu-Natal ( Daniels et al. 2019). Phylogenetically, Potamonautes mariepskoppie sp. nov. is not closely related to the five other species that occur in Mpumalanga province and can also easily be distinguished from these. Both P. unispinus and P. calcaratus have a single tooth on the anterolateral carapace margin, which is nearly spike-like in P. calcaratus . In P. unispinus the tooth on the anterolateral margin is sharp, pointed and prominent, similar to the exorbital tooth, and the species is large (CL= 49.83mm) and broad (CWW = 64.88 mm). Potamonautes unispinus is a riverine species that is widespread in southern Africa and known from South Africa, Zimbabwe and Zambia ( Stewart & Cook 1998), while P. calcaratus occurs exclusively around ephemeral pans in the Kruger National Park in South Africa, where it burrows into the clay sidewalls up to 1m ( Daniels et al. 2002a). The latter species is also found in Mozambique and Zimbabwe ( Reed & Cumberlidge 2004). Additionally, the chelipeds of P. calcaratus are highly modified and nearly flattened for burrowing. The latter species has a vaulted carapace, indicative of its semi-terrestrial mode of life (Daniels pers. obs). In Potamonautes warreni , the dentition on the anterolateral margin of the carapace varies from a single tooth to a series of five to ten teeth (Daniels 2001). Potamonautes warreni is a riverine species widespread in South Africa, Botswana and Namibia where it occurs in the Orange River and its major tributaries such as the Vaal River. Potamonautes flavusjo , a Mpumalanga Highveld endemic, is semi-terrestrial and occurs in vlei (wetland) and lake areas, where it burrows into peat soils (Daniel et al. 2014). This species has characteristic sulphur-yellow patches on its dorsal carapace surface and the ventral surface is bright yellow and the species has a highly vaulted carapace ( CH /CL = 0.61) ( Daniels et al. 2014). Potamonautes sidneyi sensu stricto is generally large-bodied (CWW> 52.4 mm) and the cephalothorax is flat ( CH /CL = 0.54), while the anterolateral margins of the carapace are heavily granulated. The species is common in large rivers, streams and swamp habitats throughout KwaZulu-Natal, Mpumalanga, Gauteng, Limpopo, the North-West and Northern Cape provinces of South Africa ( Barnard 1950; Peer et al. 2017; Daniels unpubl).
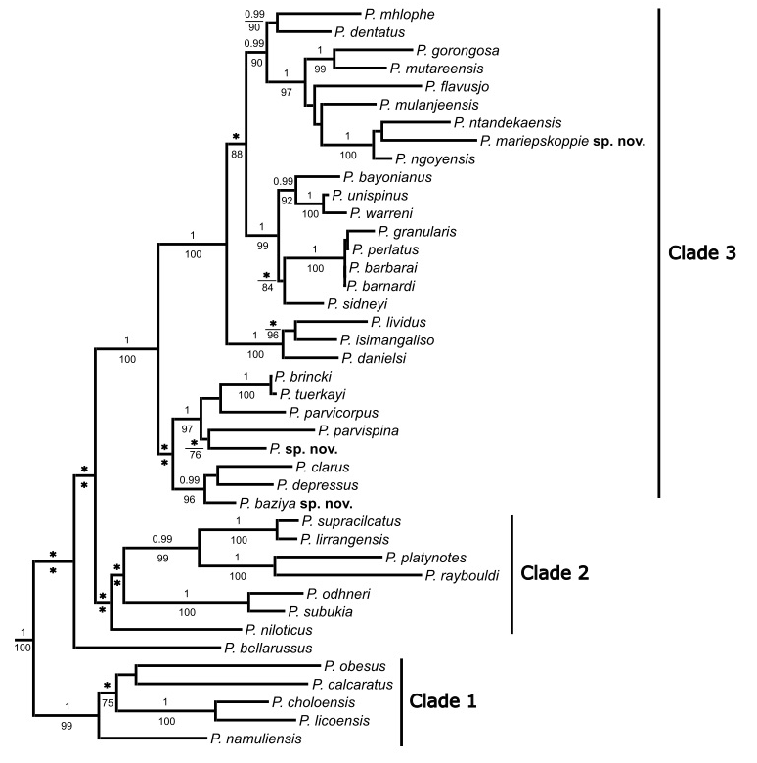
The three remaining sister species to the clade containing P. mariepskoppie sp. nov. are all found in neighboring Southern African countries. Potamonautes gorongosa occurs at Gorongosa National Park in Mozambique, P. mutariensis occurs in the Zimbabwean Highlands and P. mulanjeensis occurs on Mount Mulanje in Malawi. Phylogenetically, P. mariepskoppie sp. nov. is distantly related to the two remaining swamp forest dwelling South African freshwater crab species, P. lividus and P. isimangaliso ( Fig.1 View Fig ), although it does bear a superficial resemblance to these two species. Potamonautes lividus occurs in swamp forest patches in Eastern Cape province, at Dwesa Nature Reserve, Mazeppa Bay and Manubi State forest, and in the IOCB forests from Amatikulu Nature Reserve, Richards Bay, Empangeni, Tugela River Mouth, the University of Zululand campus, and Mapelane Nature Reserve in KwaZulu-Natal (Gouws et al. 2001; Daniels et al. 2020a). Potamonautes isimangaliso is confined to iSimangaliso Wetland Park in northern KwaZulu-Natal ( Peer et al. 2015). Potamonautes lividus was originally described from Ficus and Barringtonia dominated forests, and the carapace of the species is blue or light blue (Gouws et al. 2001). Furthermore, the cephalothorax is ovoid with no epibranchial tooth and the carapace is highly vaulted ( CH /CL = 0.64), indicative of a semi-terrestrial mode of life (Gouws et al. 2001). In P. isimangaliso , the cephalothorax is also ovoid, and the carapace is highly vaulted ( CH /CL = 0.57) and light brown, maroon, purple or brown–black in colour ( Peer et al. 2015). The species lives in ephemeral pans in sand forest where it burrows into the soil to a depth of 30–50 cm ( Peer et al. 2015). Potamonautes mariepskoppie sp. nov., is distantly related to the undescribed species from Hogsback and the four mountain-living freshwater crab species from the Cape Fold Mountains ( P. brincki , P. parvicorpus , P. parvispina and P. tuerkayi ), and the three Great Drakensberg Escarpment species ( P. clarus , P. depressus and P. baziya sp. nov).
Taxonomic note
H. Milne-Edwards (1853) described Thelphusa inflata from Durban, KwaZulu-Natal (formerly Port of Natal, Natal province). Barnard (1950) was of the opinion that Potamonautes inflatus is a variant of P. perlatus . In addition to Durban, Barnard (1950) also listed P. inflatus as being present at Belfast, Haenertsburg and Mariepskop in the former Transvaal province (notably, the last two localities are also where P. mariepskoppie sp. nov. occurs). Bott (1955) recognized P. inflatus as a junior subjective synonym of P. depressus , noting that the “type” is unknown or lost. Our phylogenetic results refute a close relationship between P. mariepskoppie sp. nov. and P. depressus . A search of the digital records of the Muséum national d’Histoire naturelle, Paris, France failed to identify the type specimen of P. inflatus , suggesting the holotype is lost. Similarly, an exhaustive search of the South African Museum of Natural History, Cape Town, failed to retrieve the material from Belfast, Mariepskop and Haenertsburg that Barnard (1950) assigned to P. inflatus . Researchers consider P. inflatus as an invalid taxon ( Stewart et al. 1995; Gouws & Stewart, 2001; Ng et al. 2008).A recent redescription of P. sidneyi suggests that the type material was likely originally collected from the region of Port Natal ( Peer et al. 2017). The original description of P. inflatus is inadequate to differentiate it from P. sidneyi . Considering the absence of type material for P. inflatus and its confusing taxonomic status, the species name a nomen nudum. In recent years, six freshwater crab species, P. dentatus , P. lividus , P. isimangaliso , P. danielsi , P. ntendekaensis and P. ngoyensis , have been described from KwaZulu-Natal. Apart from P. sidneyi , no other species occurs in close geographic proximity to Durban (Port Natal) ( Stewart et al. 1995; Peer et al. 2015, 2017; Daniels et al. 2019). The specimens from Mariepskop are recognized as P. mariepskoppie sp. nov., rather than P. inflatus , to limit any potential future confusion.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
SuperFamily |
Potamoidea |
|
Family |
|
|
SubFamily |
Potamonautinae |
|
Genus |