
Geodia phlegraei, , Koltun, 1964
|
publication ID |
https://doi.org/10.1111/zoj.12056 |
|
publication LSID |
lsid:zoobank.org:pub:5CFF222F-0C8D-4FA8-9388-D0C77213710E |
|
persistent identifier |
https://treatment.plazi.org/id/122687EB-FFF5-2140-FC0A-CCD36B21BB64 |
|
treatment provided by |
Marcus (2021-08-27 07:20:11, last updated by Plazi 2023-11-04 23:23:20) |
|
scientific name |
Geodia phlegraei |
| status |
|
GEODIA PHLEGRAEI View in CoL ( SOLLAS, 1880b)
GEODIINAE P PHLEGRAEI (PhyloCode SPECIES NAME)
( FIGS 18–20 View Figure 18 View Figure 19 View Figure 20 , 23 View Figure 23 , TABLE 5)
Isops phlegraei, Sollas, 1880b: p. 397 View in CoL ; 1888: p. 267; von Lendenfeld, 1903: p. 95; Hentschel, 1929: p. 919; Burton, 1930: p. 498; Vosmaer, 1933: p. 141; Alander, 1942: p. 73; Warén & Klitgaard, 1991: p. 52; Klitgaard, 1995: p. 2; Reitner & Hoffmann, 2003: table 1; Klitgaard & Tendal, 2004: p. 57; Cárdenas et al., 2010: p. 91.
Synops pyriformis, Vosmaer, 1882: p. 20 View in CoL ; 1885: p. 10 (synonymy by Burton, 1930).
Isops sphaeroides, Vosmaer, 1882: p. 13 View in CoL (synonymy by Burton, 1930).
Isops pallida, Vosmaer, 1882: p. 16 View in CoL (in part); Rezvoj, 1928: table 1 (synonymy by Burton, 1930).
Isops pyriformis, von Lendenfeld, 1903: p. 95 View in CoL .
Geodia phlegraei, Koltun, 1964 View in CoL (in part): p. 148; Koltun, 1966 (in part): p. 55; Cárdenas et al., 2011 (in part): table S1; Murillo et al., 2012 (in part): p. 842; Schöttner et al., 2013: p. 2.
Not:
Isops pyriformis, Hentschel, 1929: p. 865 View in CoL (= G. parva View in CoL ).
Geodia cf. phlegraei, Wagoner et al., 1989: p. 2344 View in CoL (= G. parva View in CoL ).
Isops phlegraei pyriformis, Klitgaard & Tendal, 2004: p. 57 View in CoL (= G. parva View in CoL ).
Isops phlegraei, van Soest et al., 2007: p. 129 View in CoL (= G. cf. nodastrella View in CoL ).
Geodia phlegraei, Koltun, 1966 View in CoL (in part): p. 55; Cárdenas et al., 2011 (in part): table S1; Murillo et al., 2012 (in part): p. 842 (= G. parva View in CoL ).
Misidentifications:
Pachymatisma johnstonia, Hansen, 1885: p. 17 View in CoL .
Cydonium mülleri, Breitfuss, 1912: p. 62 (according to Koltun, 1966).
Geodia cydonium, Gorbunov, 1946: p. 37 View in CoL (according to Koltun, 1966).
Type locality and type material examined
Isops phlegraei, Korsfjord near Bergen GoogleMaps , Norway. 60°10′N, 05°10′E, 330 m. Collected by Rev. A. M. Norman in 1878, BNHM 1910.1 View Materials .1.840.
Synops pyriformis , near Hammerfest , Norway, 71°12′5″N, 20°30′5″E, 247 m, Willem Barents Exp. 1878–79, RMNH Por. 660 (wet specimen); MNHN- DN23, Norman Collection, spicule preparation and section; BMNH 10.1.1.1147–1148, Norman Collection, spicule preparation and section (not seen) GoogleMaps .
Isops sphaeroides , near Hammerfest, Norway, 71°12′5″N, 20°30′5″E, 247 m, Willem Barents Exp. 1878–79, MNHN-DN24 , syntype 1, Norman Collection, spicule preparation and section ; RMNH, Vosmaer slide collection, box number 37, syntype 2, two spicule preparations and one stained thin section ; BMNH 10.1.1.1158–1159, Norman Collection, two slides (not seen).
Isops pallida , near Hammerfest, Norway, 71°12′5″N, 20°30′5″E, 247 m, Willem Barents Exp. 1878–79, RMNH Por. 652, wet specimen (seen only from pictures); RMNH, Vosmaer slide collection, box number 37, two spicule preparations with number 62.
External morphology and cortex: More or less spherical, the largest specimens somewhat flattened on the top, cup-shaped, generally up to 20 cm in diameter, and 15 cm high ( Fig. 18A, B, E View Figure 18 ); our largest specimen measures 43 cm in maximum dimension. Juveniles are spherical ( Fig. 18C View Figure 18 ). Specimens growing on vertical cliffs (i.e. fjords) can be flattened or more irregular ( Fig. 18D View Figure 18 ). Lower part sometimes formed as a base, with root-like outgrowths ( Fig. 19B View Figure 19 ). The colour alive usually varies from whitish grey to light brown, to slightly rose. One shallow fjord specimen (PC111, shallower than 175 m) was faintly purple in ethanol, like shallow G. barretti and Pachymatisma normani Sollas, 1888 (a NEA boreal Geodiidae ) ( Cárdenas & Rapp, 2013). Other specimens fixed in ethanol seemed to be reddish, although this staining comes from the epibiont yellow sponge Hexadella dedritifera Topsent, 1913 ( Fig. 18A, F View Figure 18 ) which turns dark red during the fixation. Many specimens are very hispid on the sides, but never on the top surface ( Fig. 18A–C View Figure 18 ). The fur can be up to 10 mm long, but is not a regularly occurring feature in dredged specimens as it may be easily lost during the collection. The cortex ( Fig. 18I View Figure 18 ) is thin to fairly thick (0.7–2 mm thick); it is usually difficult to cut and tends to break in pieces. Many epibionts (especially sponges) are present in this hispid part. Uniporal oscular openings are up to 1 mm in diameter, and are found mainly on the upper surface ( Fig. 18A– F View Figure 18 ). Oscular openings may be at the tip of thick conical elevations, more or less pointy, which can be lighter-coloured ( Fig. 18G View Figure 18 ) (but not always) and which gave the name to this species: Campi phlegraei is a local name of the Naples volcanic area ( Sollas, 1880b). Uniporal pores (c. 300–400 Mm in diameter) ( Fig. 18H–-J View Figure 18 ) are scattered on the sides and partly on the underside of the body. Pores are usually not elevated but can also be surrounded by a white margin.
Description of type material: The holotype of G. phlegraei is a small subspherical specimen (diameter: 2.5 cm) cut up into four parts ( Fig. 19A View Figure 19 ); it has conical-shaped oscules. There is also one BMNH slide
Means are in bold; other values are ranges; N = 30 unless stated otherwise in parentheses, or unless measurements come from other studies. A dash indicates that this measurement is not given in the literature. n.f., not found; n.o., not observed in the specimen in our possession. N, Norway; S, Sweden.
of the type but it is damaged and the embedding medium has blackened. Figure 20 View Figure 20 shows SEM pictures of the spicules from the holotype. The type of S. pyriformis is a medium-sized specimen (length: 10 cm, width: 8 cm) cut into five pieces. The main piece ( Fig. 19B View Figure 19 ) is the elongated cup-shaped half represented by Vosmaer (1882: plate IV). We have only seen pictures of the wet specimen of the lectotype (specimen a) of I. pallida ( Fig. 19C, D View Figure 19 ): the pearshaped specimen is cut into two fragments (one-half and one-third of a single specimen). The half fragment is about 6 ¥ 4 cm with a thick cortex (1–2 mm thick), uniporal oscules, and uniporal pores. There are also two spicule preparations of the lectotype (slide 62: a) of I. pallida ( Fig. 4A View Figure 4 ). One slide is broken so the label is gone but the spicules are identical to those of slide 62.
Spicules ( Fig. 20 View Figure 20 , Table 5): Megascleres: (a) oxeas, length: 1173–7600 Mm; width: 10–100 Mm. (b) Orthotriaenes, rare dichotriaenes, rhabdome length: 586– 6655 Mm; width: 12–150 Mm; orthotriaene clad length: 80–1125 Mm; protoclad length: 220–250 Mm; deuteroclad length: 100–250 Mm. (c) Anatriaenes, rare (some clads were dichotomized in the type of I. sphaeroides ) rhabdome length: up to 11 mm; width: 8–25 Mm; clad length: 30–130 Mm (minimum according to Koltun, 1966). (d) Protriaenes, very rare, rhabdome length: 11 600–12 750 Mm; width: 25–30 Mm; clad length: 190–200 Mm. Microscleres: (e) sterrasters, subspherical (NEA specimens) or spherical (in some NWA specimens), length: 82–144 Mm; width: 70–124 Mm; thickness: 65–80 Mm; hilum: 12–15 Mm. Rosettes are made of 4–12 smooth rays; rosette diameter: 6–8 Mm. (f) Spherasters with spiny conical actines (more rarely with blunt ends), 8–26 Mm in diameter. (g) Oxyasters, smooth (rough actines were rarely observed in very large oxyasters), 10–70 Mm in diameter (maximum is from measurements of the type by Sollas, 1888).
DNA barcodes: GenBank accession nos. EU442196 View Materials , HM 592701 View Materials (Folmer COI). We have sequenced COI from specimens from Spitsbergen (1), western and northern Norway (10), Mingulay Reef (1), and Orphan Knoll (1): the Folmer COI is identical in all these specimens. No. KC481222 View Materials (18S), obtained from ZMBN 77929 View Materials (Korsfjord, Norway). 18S of ZMBN 89719 View Materials (Spitsbergen) was also sequenced: no variation was observed .
Distribution ( Fig. 23 View Figure 23 ): Geodia phlegraei has a boreal distribution and seems to avoid arctic waters; it can be found from 40 m (Trondheimsfjord) to 3000 m (Orphan Knoll). It is commonly found at depths from 100–300 m (Norwegian continental shelf) to 725 m (Faeroes), at temperatures of 0.3 °C (BIOICE, st. 2926) to 7.9 °C (BIOFAR, st. 297). It has also been recorded by divers in Norwegian fjords at shallower depths: for example, it has been photographed by A. Salesjö in the Trondheimsfjord at only 40–50 m depth (http:// www.anderssalesjo.com/?id=3306&lang=42, accessed 7 May 2013). We also identified it in material collected in Mingulay Reef (western Scotland) at 128–139 m depth. In Mingulay reef (R. van Soest, pers. comm.) and Norwegian fjords at shallow depths, temperatures can reach 10.5 °C. The specimen identified as G. phlegraei from Rockall Bank ( van Soest et al., 2007) was actually a Geodia cf. nodastrella . Interestingly, all but one of the NWA specimens from the Flemish Cap that we examined (a total of eight specimens) collected during NEREIDA 2009–10 and originally identified as G. phlegraei were in fact G. parva . The only specimen from the Flemish Cap that we identified as G. phlegraei (DR24-69d = UPSZMC 78280) has an external morphology similar to G. phlegraei and large elongated sterrasters (c. 132–136 Mm in diameter). Unfortunately, we could not get a COI sequence from it to confirm this identification. However, we did get a COI sequence for R1340-04 (= UPSZMC 78308) confirming it was G. phlegraei : it has large spherical sterrasters (88–107 Mm in length) and it was collected at 3000 m depth at Orphan Knoll, where the temperature was 2.4 °C ( Fig. 18F View Figure 18 shows R1341-18 collected at 2347 m in the same area). The most western specimen of G. phlegraei found is from western Greenland (‘Shinkai Maru’, st. 32, 64°13.5′N, 54°42.1′W. 970 m) ( Fig. 18G, H View Figure 18 ), but this material was not suited for molecular studies.
Biology: When he described this species, Sollas (1880) immediately noticed that it was ‘covered by various foreign bodies’. Indeed, G. phlegraei is the boreoarctic Geodia which is most often found covered with epifauna, especially sponges and including individuals of its own species. Here are a few sponges found on G. phlegraei : Craniella sp. , Cyamon spinispinosum ( Topsent, 1904) , Hexadella dedritifera (mis-identified as Aplysilla sulphurea in Klitgaard (1995)), Leucandra spp. , Lissodendoryx (L.) fragilis Fristedt, 1885 , Polymastia grimaldii ( Topsent, 1913) , Stelletta normani , Ute gladiata Borojevic, 1966 , etc. G. phlegraei can settle on other sponges as well such as other large Astrophorina [ S. normani , Stryphnus fortis ( Vosmaer, 1885) ]. The predator chiton H. nagelfar and the parasitic foraminiferan H. sarcophaga have been found living on G. phlegraei ( Warén & Klitgaard, 1991; Cedhagen, 1994; Todt et al., 2009). More associated fauna has been investigated by Klitgaard (1995). The chemistry (elemental analysis, amino acids, sterols, and quaternary ammonium compounds) has been investigated by Hougaard et al. (1991a, b).
The associated microsymbionts of G. phlegraei collected in the Sula Ridge reef ( Norway) have been studied ( Graeber et al., 2004; Dieckmann et al., 2005) and led to the isolation and description of a new gammaproteobacterium ( Oceanospirillales group): Spongiispira norvegica ( Kaesler et al., 2008) .
We observed on the type section of I. sphaeroides made by Vosmaer many subglobular oocytes without pseudopodes. This specimen has been collected in the Barents Sea on 2 July 1879.
In the NEA, G. phlegraei can easily be confused with another Geodiidae , Pachymatisma normani , which also has raised white-rimmed uniporal oscules, but P. normani has microrhabds in the cortex, instead of spheroxyasters. Geodia phlegraei can also be easily confused with its sister species G. parva , and in that case only spicule and genetic characters can differentiate them (cf. below).
Distinctive characters: External morphology: round to sometimes cup-shaped, with only uniporal openings (i.e. no sieve). The numerous small oscules on the top of specimen, each with a whitish rim making it look like a little wart. Often overgrown with other sponges, hydrozoa, bryozoa, etc. Spicules: large spherasters, smooth oxyasters along with fairly large sterrasters (70–144 Mm).
Remarks: Choanosomal oxyasters can sometimes be separated into two size categories (10–25 and 45–60 Mm) but in some specimens we have more of a continuum of sizes so we decided to treat oxyasters as one category ( Table 5). Koltun (1966) notes the presence of rare small slightly curved oxeas (230–420 ¥ 8–10 Mm); we never observed those and wonder whether these could have been contamination. Dichotriaenes have been reported by Koltun (1966) and Vosmaer (1882), but it should be stressed here that they are fairly rare as we only found a few (notably in the holotype). Anatriaenes are rare and we report for the first time the presence of protriaenes in this species (found in the type of I. sphaeroides ), but they seem to be very rare.
Geodia phlegraei View in CoL is the type species of Isops Sollas, 1880b View in CoL , a genus synonymized with Geodia View in CoL based on molecular results ( Cárdenas et al., 2010). Synops pyriformis View in CoL is the type species of Synops View in CoL , a genus later synonymized with Isops ( Sollas, 1889) View in CoL . Vosmaer’s (1882) species ( Isops sphaeroides View in CoL , Isops pallida View in CoL , Synops pyriformis View in CoL ) were all described from the same station near Hammerfest (71°12′5″N, 20°30′5″E) and collected on the same day (2 July 1879). Burton (1930: 498) writes: ‘Examinations of the preparations in the Norman Collection, made from the type-specimens of these species, show conclusively that Geodia parva Hansen View in CoL , Isops pallida Vosmaer View in CoL , I. sphaeroides Vosmaer View in CoL , and Synops pyriformis Vosmaer View in CoL are synonymous with Isops phlegraei Sollas. View in CoL ’ We stated earlier that Burton did not look at the type of I. pallida View in CoL , and that a lectotype ( Fig. 19C, D View Figure 19 ) was designated to keep the synonymy with G. phlegraei View in CoL . On the other hand, Burton did look at the BMNH type slides of I. sphaeroides View in CoL and S. pyriformis View in CoL .
Isops sphaeroides View in CoL was described from two globular syntypes (both presumably lost, R. van Soest & N. de Voogd, pers. comm.) : one larger specimen was hispid, whereas the smaller was not. All type slides we examined were identified as G. phlegraei View in CoL ( Table 5). However, we found differences between the MNHN type slides ( Norman Collection ) and those from the Vosmaer slide collection, namely differences in cortex thickness and sterraster sizes ( Table 5), so these may represent the two syntypes. The Vosmaer slides include a section from half of a smooth specimen with an estimated total diameter of 2–2.5 cm, which must be the smaller syntype . To conclude, we presume that the Vosmaer slides correspond to the smaller syntype so the MNHN slides correspond to the larger syntype .
We also compared type specimen and type slides of S. pyriformis with our material. The cortex thickness of S. pyriformis (1.1–1.3 mm) agrees well with G. phlegraei ( Table 5). Spicule measurements also match those of G. phlegraei ( Table 5). Finally, observation of the external morphology ( Fig. 19B View Figure 19 ) and new spicule preparations from the wet type of S. pyriformis further confirmed this. All in all, we therefore follow Burton (1930) and confirm the synonymy for S. pyriformis , I. pallida , and I. sphaeroides .
Koltun (1966) had already underlined the morphological variability of G. phlegraei . He notably states that in deeper and colder waters the morphology of G. phlegraei is somewhat different. Klitgaard & Tendal (2004) also noticed this arctic water morphotype and considered it as a subspecies of G. phlegraei : G. phlegraei pyriformis . After having examined many specimens from the whole boreo-arctic area, we confirm the existence of different morphotypes, and, after incorporating additional morphological data as well as molecular data, it was decided that the arctic morphotype represented a valid species which had in fact been previously described under the name Geodia parva Hansen, 1885 , before being synonymized with G. phlegraei ( Burton, 1930) . Below, we resurrect and redescribe G. parva .
Alander H. 1942. Sponges from the Swedish West-Coast and adjacent waters. PhD Thesis, Lund University. Henrik Struves: Goteborg.
Borojevic R. 1966. Eponges calcaires des cotes de France. III. Discussion sur la taxonomie des Eponges Calcaires: Aphroceras ensata (Bowerbank) et Ute gladiata sp. n. Archives de Zoologie Experimentale et Generale 107: 703 - 723.
Breitfuss JS. 1912. Zur Kenntnis der Spongio-Fauna des Kola Fjords. Travaux de la Societe Imperiale des Naturalistes de St. Petersbourg, Section Zoologie 41: 61 - 80, pls I - II.
Burton M. 1930. Norwegian sponges from the Norman collection. Proceedings of the Zoological Society of London 1930: 487 - 546, pls I - II.
Cardenas P, Rapp HT, Schander C, Tendal OS. 2010. Molecular taxonomy and phylogeny of the Geodiidae (Porifera, Demospongiae, Astrophorida) - combining phylogenetic and Linnaean classification. Zoologica Scripta 39: 89 - 106.
Cardenas P, Xavier JR, Reveillaud J, Schander C, Rapp HT. 2011. Molecular phylogeny of the Astrophorida (Porifera, Demospongiae) reveals an unexpected high level of spicule homoplasy. PLoS ONE 6: e 18318.
Cardenas P, Rapp HT. 2013. Disrupted spiculogenesis in deep-water Geodiidae (Porifera, Demospongiae) growing in shallow waters. Invertebrate Biology. doi: 10.1111 / ivb. 12027.
Cedhagen T. 1994. Taxonomy and biology of Hyrrokin sarcophaga gen. et sp. n., a parasitic foraminiferan (Rosalinidae). Sarsia 79: 65 - 82.
Dieckmann R, Graeber I, Kaesler I, Szewzyk U, von Dohren H. 2005. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by intact-cell-MALDI-TOF mass spectrometry (ICM-MS). Applied Microbiology and Biotechnology 67: 539 - 548.
Fristedt K. 1885. Bidrag till Kannedomen om de vid Sveriges vestra Kust lefvande Spongiae. Kungliga Svenska vetenskapsakademiens handlingar 21: 1 - 56, pls I - IV.
Gorbunov GP. 1946. Benthonic populations of the novosibirsk shallow and central part of the Arctic Ocean. Tr. dreif. eksped. Glavsevmorputi na 1 / p ' G. Sedov' 1937 - 1940 III: 30 - 138 (in Russian).
Graeber I, Kaesler I, Dieckmann R, von Dorhen H, Szewzyk U. 2004. Sponge associated bacteria from boreal sponges. Bollettino dei musei e degli istituti biologici dell' Universita di Genova 68: 357 - 363.
Hansen GA. 1885. Spongiadae. The Norwegian North-Atlantic Expedition 1876 - 1878. Zoologi 13: 1 - 26. pls I - VII, 21 maps.
Hentschel E. 1929. Die Kiesel- und Hornschwamme des Nordlichen Eismeers. In: Romer F, Schaudinn F, Brauer A, Arndt W, eds. Fauna Arctica. Eine Zusammenstellung der arktischen Tierformen mit besonderer Berucksichtigung des Spitzbergen-Gebietes auf Grund der Ergebnisse der Deutschen Expedition in das Nordliche Eismeer im Jahre 1898. Jena: G. Fischer, 857 - 1042, pls XII - XIV.
Hougaard L, Anthoni U, Christophersen C, Larsen C, Nielsen PH. 1991 a. Characterization and quantitative estimation of quaternary ammonium compounds in marine demosponges. Comparative Biochemistry and Physiology 99 B: 469 - 472.
Kaesler I, Graeber I, Borchert MS, Pape T, Dieckmann R, von Dohren H, Nielsen P, Lurz R, Michaelis W, Szewzyk U. 2008. Spongiispira norvegica gen. nov., sp. nov., a marine bacterium isolated from the boreal sponge Isops phlegraei. International Journal of Systematic and Evolutionary Microbiology 58: 1815 - 1820.
Klitgaard AB. 1995. The fauna associated with outer shelf and upper slope sponges (Porifera, Demospongiae) at the Faroe Islands, northeastern Atlantic. Sarsia 80: 1 - 22.
Klitgaard AB, Tendal OS. 2004. Distribution and species composition of mass occurrences of large-sized sponges in the northeast Atlantic. Progress in Oceanography 61: 57 - 98.
Koltun VM. 1964. Sponges (Porifera) collected in the Greenland seas and from the region to the north of the Spitzbergen and Franz Josef Land, from expeditions of the ' F. Litke' 1955, ' Obb' 1956 and ' Lena' 1957 and 1958. Scientific results of the high- latitudes Oceanographic Expeditions to the northern part of the Greenland Sea and adjacent areas of the Arctic basin between 1955 - 1958. Trudy Arkticheskogo i antarkticheskogo Nauchno- Issledovatel'skogo Instituta 259: 143 - 166 (in Russian).
Koltun VM. 1966. Four-rayed sponges of Northern and Far Eastern seas of the USSR (order Tetraxonida). Opredeliti Faunei SSSR 90. (Zoological Institute of the Academy of Sciences of the USSR: Moscow, Leningrad): 1 - 112, pls I - XXXVIII (in Russian).
Murillo FJ, Munoz PD, Cristobo J, Rios P, Gonzalez C, Kenchington E, Serrano A. 2012. Deep-sea sponge grounds of the Flemish Cap, Flemish Pass and the Grand Banks of Newfoundland (Northwest Atlantic Ocean): Distribution and species composition. Marine Biology Research 8: 842 - 854.
Reitner J, Hoffmann F. 2003. Schwamme in Kaltwasser- Korallenriffen. Kleine Senckenberg-Reihe 45: 75 - 87.
Rezvoj P. 1928. Contribution to the fauna of Porifera in the Barents Sea. Transactions of the Institute for scientific Exploration of the North 37: 67 - 95.
Schottner S, Hoffmann F, Cardenas P, Rapp HT, Boetius A, Ramette A. 2013. Relationships between host phylogeny, host type and bacterial community diversity in cold-water coral reef sponges. PLoS ONE 8: e 55505. doi: 10.1371 / journal. pone. 0055505
Sollas WJ. 1880 b. The sponge-fauna of Norway; a Report on the Rev. A. M. Norman's Collection of Sponges from the Norwegian Coast. Annals and Magazine of Natural History 5: 396 - 409, pl. XVII.
Sollas WJ. 1888. Report on the Tetractinellida collected by H. M. S. Challenger, during the years 1873 - 1876. Report on the Scientific Results of the Voyage of H. M. S. Challenger, 1873 - 1876. Zoology 25: 1 - 458, pls I-XLIV, 451 map.
Sollas WJ. 1889. On the geodine genera, Synops, Vosm., and Sidonops. A correction. Scientific Proceedings of the Royal Dublin Society 6: 276 - 277.
Todt C, Cardenas P, Rapp HT. 2009. The chiton Hanleya nagelfar (Polyplacophora, Mollusca) and its association with sponges in the European Northern Atlantic. Marine Biology Research 5: 408 - 411.
Topsent E. 1904. Spongiaires des Acores. Resultats des campagnes scientifiques accomplies par le Prince Albert I. Monaco 25: 1 - 280, pls 281 - 218.
Topsent E. 1913. Spongiaires provenant des campagnes scientifiques de la ' Princesse Alice' dans les Mers du Nord (1898 - 1899 - 1906 - 1907). Resultats des campagnes scientifiques accomplies par le Prince Albert I. Monaco 45: 1 - 67, pls I - V.
van Soest RWM, Cleary DFR, de Kluijver MJ, Lavaleye MSS, Maier C, van Duyl FC. 2007. Sponge diversity and community composition in Irish bathyal coral reefs. Contributions to Zoology 76: 121 - 142.
von Lendenfeld R. 1903. Porifera. Tetraxonia. In: Schulze FE, ed. Das Tierreich. Berlin: Friedlander, vi - xv, 1 - 168.
Vosmaer GCJ. 1882. Report on the sponges dredged up in the Arctic Sea by the ' Willem Barents' in the years 1878 and 1879. Niederlandisches Archiv fur Zoologie Supplement 1: 1 - 58, pls I - IV.
Vosmaer GCJ. 1885. The sponges of the ' Willem Barents' Expedition 1880 and 1881. Bijdragen tot de Dierkunde 12: 1 - 47, pls I-V.
Vosmaer GCJ. 1933. The sponges of the Bay of Naples, Porifera Incalcaria. With analyses of genera and studies in the variations of species. Martinus Nijhoff: The Hague. 456 pp.
Wagoner NAV, Mudie PJ, Cole FE, Daborn G. 1989. Siliceous sponge communities, biological zonation, and Recent sea-level change on the Arctic margin: Ice Island results. Canadian Journal of Earth Sciences 26: 2341 - 2355.
Waren A, Klitgaard A. 1991. Hanleya nagelfar, a spongefeeding ecotype of H. hanleyi or a distinct species of chiton? Ophelia 34: 51 - 70.

Figure 4. A, type slides of Isops pallida Vosmaer, 1882 for specimen numbers 62 (= a, here designated as lectotype) identified as G. phlegraei, and 64 (= b) identified as G. atlantica. B, orthotriaenes and abundant anatriaenes from one of the slides 64 (= b). C and D, thick sections of the holotype of Sidonops atlantica Stephens, 1915, specimen S.R.151-27/ 364–1914. E and F, thick sections of ZMBN 77927, Korsfjord, 200–400 m depth. Scale bars: B, 400 Mm; C, E, F: 500 Mm; D, 300 Mm.

Figure 18. External morphology and thick sections of Geodia phlegraei Sollas, 1880b. A, specimen PC124 just after dredging from the Korsfjord, Norway, 200–400 m depth; specimen is 20 cm in diameter. B, specimen ZMBN 85211 just after collecting from Traenadjupet, northern Norway, 290 m depth: specimen is 20 cm in diameter. C, specimen PC68 just after dredging from the Korsfjord, Norway, 200–400 m depth; specimen is 4 cm in diameter. D, specimen PC191 just after dredging from the Korsfjord, Norway, 200–400 m depth; specimen is 20 cm in length. E, specimen from Ingolf Exp., st. 90, Denmark Strait, 1039 m depth (fixed in ethanol); specimen is 15 cm high and 8 cm in diameter. F, specimen R1341-18 in situ, collected at 2347 m depth, Orphan Knoll (50.05894, 45.60638); specimen is 20 cm in length and covered with the yellow encrusting sponge Hexadella dedritifera (Canadian DFO/ROPOS 2010). G, uniporal oscules (specimen from Shinkai Maru Exp., st. 32). H, uniporal pores (specimen from Shinkai Maru Exp., st. 32). I, thick section from ZMBN 77929 (Korsfjord, 200–400 m). J, uniporal pore with sphincter. Scale bars: G: 5 mm; H: 1 cm; I: 1 mm; J: 200 Mm.
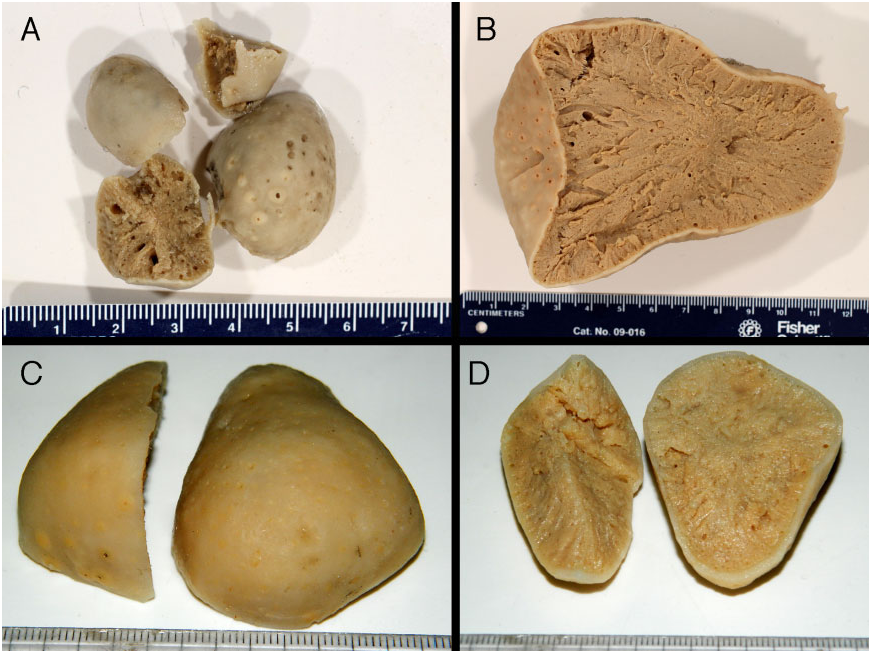
Figure 19. A, holotype of Geodia phlegraei SOLLAS, 1880b, BMNH 1910.1.1.840. B, holotype of Synops pyriformis Vosmaer, 1882, RMNH Por. 660. C, lectotype of Isops pallida Vosmaer, 1882, RMNH Por. 652, external appearance (picture: R. van Soest). D, lectotype of I. pallida, RMNH Por. 652, internal appearance (picture: R. van Soest).

Figure 20. Spicules of the holotype (BMNH 1910.1.1.840) of Geodia phlegraei Sollas, 1880b. A, oxyasters. B, spherasters. A and B have the same scale. C, orthotriaene. D, sterrasters. E, close-up on the hilum of a sterraster. Note the smooth rosettes. Scale bar: 3 Mm.

Figure 23. Distribution of Geodia phlegraei Sollas, 1880b (dark dots) and Geodia parva Hansen, 1885 (light dots). White dots represent specimens of G. phlegraei or G. parva but that we could not assign to the proper species for various reasons (map made with GeoMapApp, http://www.geomapapp.org). T, type locality.
| RMNH |
National Museum of Natural History, Naturalis |
| HM |
Hastings Museum |
| COI |
University of Coimbra Botany Department |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Geodia phlegraei
| Cárdenas, Paco, Rapp, Hans Tore, Klitgaard, Anne Birgitte, Best, Megan, Thollesson, Mikael & Tendal, Ole Secher 2013 |
Isops phlegraei
| van Soest RWM & Cleary DFR & de Kluijver MJ & Lavaleye MSS & Maier C & van Duyl FC 2007: 129 |
Isops phlegraei pyriformis, Klitgaard & Tendal, 2004 : p. 57
| Klitgaard AB & Tendal OS 2004: 57 |
Geodia cf. phlegraei
| Wagoner NAV & Mudie PJ & Cole FE & Daborn G 1989: 2344 |
Geodia cydonium, Gorbunov, 1946 : p. 37
| Gorbunov GP 1946: 37 |
Isops pyriformis, Hentschel, 1929 : p. 865
| Hentschel E 1929: 865 |
mülleri, Breitfuss, 1912 : p. 62
| Breitfuss JS 1912: 62 |
Isops pyriformis
| von Lendenfeld R 1903: 95 |
Pachymatisma johnstonia
| Hansen GA 1885: 17 |
Synops pyriformis, Vosmaer, 1882 : p. 20
| Vosmaer GCJ 1882: 20 |
Isops sphaeroides, Vosmaer, 1882 : p. 13
| Vosmaer GCJ 1882: 13 |
Isops pallida, Vosmaer, 1882 : p. 16
| Vosmaer GCJ 1882: 16 |
Isops phlegraei, Sollas, 1880b : p. 397
| Cardenas P & Rapp HT & Schander C & Tendal OS 2010: 91 |
| Klitgaard AB & Tendal OS 2004: 57 |
| Klitgaard AB 1995: 2 |
| Waren A & Klitgaard A 1991: 52 |
| Alander H 1942: 73 |
| Vosmaer GCJ 1933: 141 |
| Burton M 1930: 498 |
| Hentschel E 1929: 919 |
| von Lendenfeld R 1903: 95 |
| Sollas WJ 1880: 397 |