
Geodia macandrewii, BOWERBANK, 1858
|
publication ID |
https://doi.org/ 10.1111/zoj.12056 |
|
publication LSID |
lsid:zoobank.org:pub:5CFF222F-0C8D-4FA8-9388-D0C77213710E |
|
persistent identifier |
https://treatment.plazi.org/id/122687EB-FFCF-214B-FF76-CDE16A69B9B3 |
|
treatment provided by |
Marcus |
|
scientific name |
Geodia macandrewii |
| status |
|
GEODIA MACANDREWII BOWERBANK, 1858 View in CoL
CYDONIUM P MACANDREWII (PhyloCode SPECIES NAME)
( FIGS 15–17 View Figure 15 View Figure 16 View Figure 17 , TABLE 4)
Geodia macandrewii, Bowerbank, 1858: p. 284 View in CoL ; 1872a: p. 196; Schmidt, 1866: p. 11 –12; Koltun, 1966: p. 50; Barthel et al., 1991: p. 40; Hougaard et al., 1991a: p. 225, b: p. 469; Warén & Klitgaard, 1991: p. 52; Ereskovsky, 1993: p. 23; Klitgaard 1995: p. 2; Reitner & Hoffmann, 2003: table 1; Murillo et al., 2012: p. 842; Schöttner et al., 2013: p. 2. Geodia macandrewia (misspelling), Cedhagen, 1994: p. 67. Geodia macandrewi (misspelling), Klitgaard & Tendal, 2004: p. 57; Cárdenas et al., 2010: p. 91; Cárdenas et al., 2011: table S1.
Cydonium normani, Sollas, 1888: p. 263 View in CoL ; Norman, 1893: p. 347; Vosmaer, 1933: p. 102 (synonymy by this study).
Synops macandrewii, Sollas, 1888: p. 265 View in CoL .
Sidonops macandrewii, Sollas, 1889: p. 277 ; von Lendenfeld, 1903: p. 100. Sidonops macandrewi (misspelling), Brøndsted, 1932: p. 5.
Geodia normani, Burton, 1930: p. 490 View in CoL (synonymy by this study).
Misidentification:
Geodia megastrella, Kingston et al., 1979: p. 528 View in CoL .
Type locality and deposition of holotype: Collected by Robert McAndrew. Vikna Island (formerly called Vigten or Vikten Island), North Trøndelag, Norway, 183 m. The holotype specimen was not found (E. Sherlock, BNHM, pers. comm.); only type slides were examined: BNHM 1877.5 View Materials .21.1396 (surface and two spicule preparations) and BNHM 1877.5 View Materials .21.1398 (one section, one spicule preparation) .
External morphology and cortex: Young specimens are spherical ( Fig. 15E View Figure 15 ). The regular spherical body form persists during growth until the body reaches a size of about 10 cm in diameter; from that on the diameter is larger than the height, and our largest specimens are somewhat flattened, measuring 35–42 cm in maximum diameter and 20–24 cm in height ( Fig. 15A– C View Figure 15 ). Some mid-sized specimens from eastern Greenland and the Flemish Cap ( Fig. 15D View Figure 15 ) are unusually flat, the diameter measuring more than twice the height. The colour alive is whitish yellow to light grey; whitish in ethanol. The choanosome is yellowish in live specimens ( Fig. 15B View Figure 15 ); whitish in ethanol. Small specimens have smooth surfaces; larger ones develop a fur of long spicules, essentially on the sides ( Fig. 15A View Figure 15 ), which is frequently damaged during the catching procedure, so that large surface areas appear smooth. Cribriporal oscules ( Fig. 15F View Figure 15 ) are regularly scattered over the upper surface; each sieve is 0.5–1 mm in diameter. Cribriporal pores ( Fig. 15G View Figure 15 ) are scattered over the sides of the body; sieves are also 0.5–1 mm in diameter. The sterraster layer is very strong and tough, and usually 1–2 mm thick (occasionally up to 4 mm thick) ( Fig. 15H, I View Figure 15 ). The ectocortex with spheroxyasters and microxeas is very thin (45–100 Mm thick) compared with the endocortex ( Fig. 15H, I View Figure 15 ).
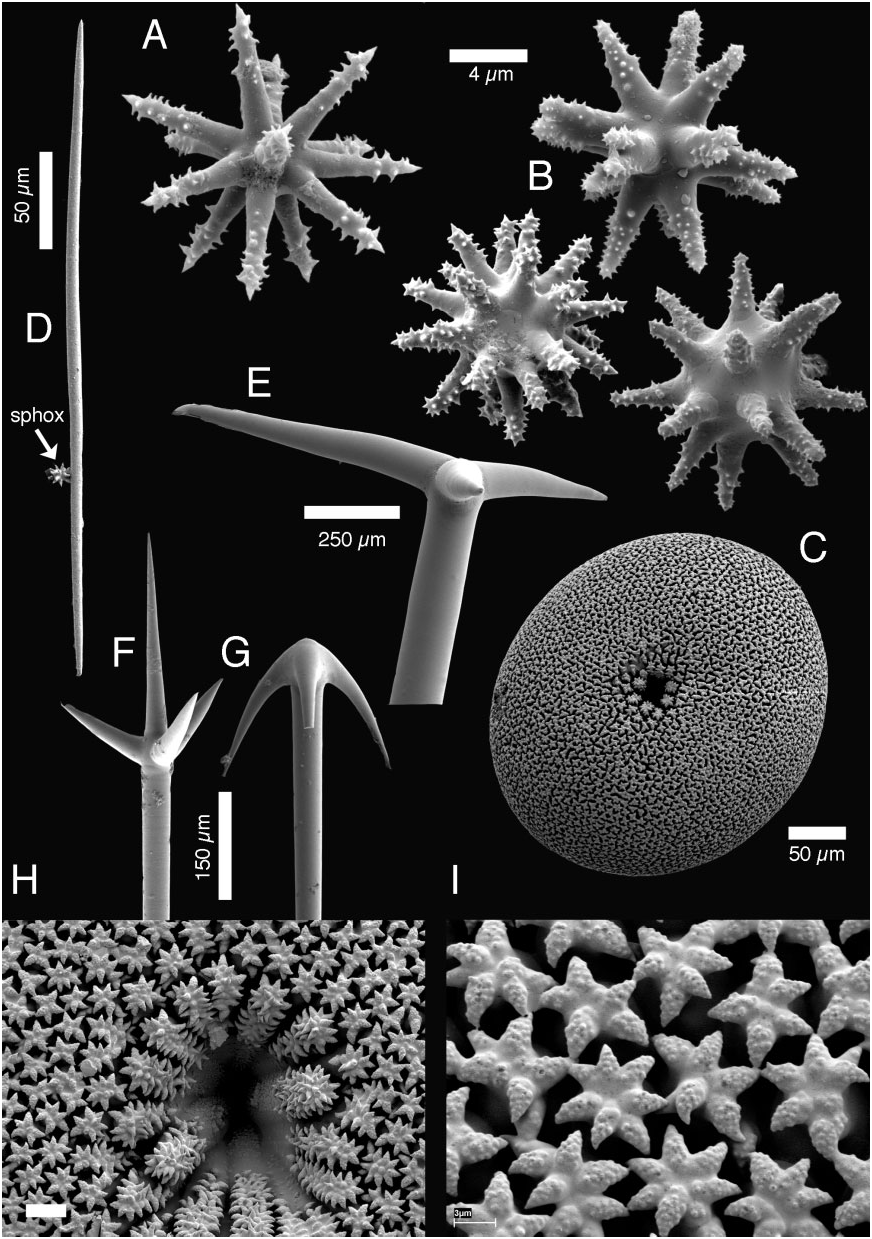
Spicules ( Fig. 16 View Figure 16 , Table 4): Megascleres: (a) oxeas, straight or bent, length: up to 14 mm; width: 5–106 Mm. (b) Microxeas, straight or slightly bent, rarely centrotylote, length: 220–445 Mm; width: 3–13 Mm. (c) Ortho- to dichotriaenes, orthotriaenes are more common, straight rhabdome, rhabdome length: 1650–9625 Mm (maximum from Brøndsted, 1932); width: 40–165 Mm. Clads can often end with a small downward bend, orthotriaene clad length: 210– 1125 Mm; protoclad length: 220–500 Mm; deuteroclad length: 70–450 Mm. (d) Anatriaenes, common swelling on top of the cladome, rhabdome length up to 22 mm; width: 3.5–63 Mm; clad length: 24–285 Mm. (e) Meso/ protriaenes, rhabdome length: up to 12 mm; width: 5–79 Mm; clad length: 32–330 Mm; central clad length: 49–685 Mm. Microscleres: (f) sterrasters, spherical to subspherical, 124–360 Mm in diameter; thickness: 128–230 Mm; hilum diameter: 20–30 Mm. Rosettes are made of 5–7 warty rays; rosette diameter: 6–10 Mm. (g) Spheroxyasters, rough actines (difficult to see with the optical microscope), with centrum more or less developed, diameter: 4–18 Mm. (h) Oxyasters, thin rough actines, diameter: 10–88 Mm.
DNA barcodes: GenBank accession nos. EU442198 View Materials , HM 592689 View Materials , HM 592696 View Materials (Folmer COI): we have sequenced COI from specimens from western and northern Norway (4), Spitsbergen (2), Davis Strait (1), and Flemish Cap (1): the Folmer COI is identical in all these specimens. No. EU552082 View Materials (28S, C1-D2 domains): we have sequenced 28S from the Bergen area (2), and Spitsbergen (1), 1-bp difference was observed between the two Bergen specimens. No. KC481225 View Materials (18S), obtained from ZMBN 89717 View Materials (Spitsbergen). Two specimens from Spitsbergen sequenced: no variation was observed .
Distribution ( Fig. 17 View Figure 17 ): We characterize the species as northern boreal, with the ability to invade some neighbouring cold-water areas, but not penetrating into real Arctic conditions. It has been recorded at depths from 157 m (Trondheimsfjord) to 1900 m (eastern Greenland). The temperature range is –0.82 °C (north of the Faroe Islands, BIOFAR st. 901) to 8.3 °C (south-west of the Faroe Islands, BIOFAR st. 69). Most eastern records are from between 230 and 400 m, at temperatures of 5–8 °C. Occurrences at temperatures below 2 °C were essentially found in the Denmark Strait (282–467 m depth). The records on the southern flanks of the Bill Bailey and Faroe Banks came from rather deep water, 1140 and 650 m, respectively. They hardly represent the southern distribution limit of the species but could indicate that off the Shetland Isles and Scotland it can be expected to occur on the upper slope. Brøndsted (1932) and Koltun (1966) mentioned the Shetlands as part of the distribution area; although this is to be expected, there is so far no proof, the record being a mistake by Brøndsted for the Norwegian record of the type specimen. Koltun (1966) mentioned occurrences in the south-western Barents Sea and the Denmark Strait, but gave no detailed information.
Biology: We found no indications of asexual reproduction. The predator chiton H. nagelfar and the parasitic foraminiferan H. sarcophaga have been found living on G. macandrewii ( Warén & Klitgaard, 1991; Cedhagen, 1994; Todt et al., 2009). More associated fauna has been investigated by Klitgaard (1995). The chemistry (elemental analysis, amino acids, sterols, and quaternary ammonium compounds) has been investigated ( Kingston et al., 1979; Hougaard et al., 1991a, b). Note that G. macandrewii off the Labrador coast ( Canada) and from the Faroe Islands have very similar sterol composition ( Kingston et al., 1979; Hougaard et al., 1991b).
Distinctive characters: External morphology: the regularly round, almost spherical form with no conspicuous openings, the hard consistency, the uniform distribution on the top side of cribriporal oscules, and the very thick cortex. Spicules: very large sterrasters (124–360 Mm in diameter).
Remarks: It was named after Robert MacAndrew who collected this species. Spicule measurements of this species are scarce apart from Sollas (1888), Brøndsted (1932), and Koltun (1966). Although this is a fairly common species in some areas, it is rarely mentioned in the literature compared with G. barretti or G. phlegraei , perhaps because it is often confused Means are in bold; other values are ranges; N = 30 unless stated otherwise in parentheses, or unless measurements come from other studies. A dash indicates that this measurement is not given in the literature. n.f., not found.
with these species. In spicule preparations, it can be difficult to separate the largest ectocortical spheroxyasters from the smallest choanosomal oxyasters as there is no clear transition from one form to the other, so we recommend measuring them on a section, to ensure not to mix both categories (this was done for the holotype and ZMBN 77924). There is some variation within oxyasters: small (up to 25–28 Mm) with thin actines (1–2 Mm thick) usually in specimens from shallower depths (183–600 m), to very large oxyasters (40–88 Mm) with thick actines (4–7 Mm thick) in specimens living deeper than 1000 m. The large sizes are not mentioned by Koltun (1966). The large dichotriaenes with atypical forward orientated cladomes represented by Bowerbank (1872a: plate X, fig. 4) were indeed observed on the type slide number ‘Bk.1398’. However, in our opinion, they result from a contamination from another Astrophorina (probably Stelletta normani Sollas, 1880a , a common species on the Norwegian coast at these same depths).
Molecular phylogenetic studies suggest that G. macandrewii belongs to Cydoniump along with G. cydonium , G. conchilega , and Geodia papyracea Hechtel, 1965 ( Fig. 2 View Figure 2 ). G. macandrewii shares the possession of identical cribriporal oscules and pores with G. cydonium and G. conchilega ; G. papyracea , by contrast, has uniporal oscules ( Cárdenas et al., 2009). We could nonetheless suggest that the association of cribriporal oscules and oscules could be an autapomorphy of the clade, and that the cribriporal oscules were later modified and lost in G. papyracea . These four species also share spiny euasters but spiny euasters are also present in the neighbouring Geodia clades (Depressiogeodiap and Geodiap) ( Fig. 2 View Figure 2 ). Geodia macandrewii , G. cydonium , and G. papyracea share orthotriaenes but G. conchilega has dichotriaenes. So additional sampling is needed to better understand the Cydoniump clade and find morphological apomorphies that would support it.
No spicule differences were observed between NWA and NEA specimens. Sterrasters are usually subspherical but we noted that one specimen from Davis Strait ( UPSZMC 78255 ) had some atypically shaped sterrasters, most of them ‘lemon-shaped’, but other specimens from the same locality had ‘normal’ sterrasters .
Geodia normani ( Sollas, 1888) View in CoL is known only from the original description. The single specimen was collected by Rev. Canon A. M. Norman at Vikna Island, which is also the type locality for G. macandrewii View in CoL . Burton (1930) worked on the Norman collection, but indicated that the single specimen was not at his disposal. Accordingly the specimen may have been untraceable even at that time. New searches in the BMNH collection for the present study have not been successful (E. Sherlock, BMNH, pers. comm., September 2012) so the type is presumably lost. We may suppose that Sollas (1888) did not examine the type specimen of G. macandrewii View in CoL while describing G. normani View in CoL , and misinterpreted Bowerbank’s description of G. macandrewii View in CoL . Indeed, Sollas (1888) moved G. macandrewii View in CoL to the genus Synops Vosmaer, 1882 View in CoL and therefore did not think of comparing it with G. normani View in CoL that he had placed in Cydonium Fleming, 1828 (because of the presence of cribriporal pores and oscules). However, the description of G. normani View in CoL , notably concerning the fairly thick cortex, the cribriporal openings, and the spicule morphologies (large spherical sterrasters, common promesotriaenes, and anatriaenes with swollen cladomes), matches that of G. macandrewii View in CoL . Geodia normani View in CoL only differs from G. macandrewii View in CoL by the absence of microxeas and we can suppose that these may have been overlooked or Sollas more likely thought they were a contamination. To conclude, because G. normani View in CoL and G. macandrewii View in CoL come from the same type locality and share so many characters (external morphology and spicules), we formally propose to consider G. normani View in CoL as a junior synonym of G. macandrewii View in CoL .
We examined slides (74-8-27.1) made by H. M. Reiswig from material published under the name Geodia megastrella and collected off the Labrador coast (53°24′50″N, 52°15′00″W) at 732 m depth ( Kingston et al., 1979). We observed a thick cortex (2 mm thick), orthotriaenes, common ana/protriaenes, very large sterrasters (up to 360 Mm), and spiny oxyasters with thin actines (10–30 Mm). This leaves no doubt that this is in fact G. macandrewii .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Geodia macandrewii
| Cárdenas, Paco, Rapp, Hans Tore, Klitgaard, Anne Birgitte, Best, Megan, Thollesson, Mikael & Tendal, Ole Secher 2013 |
Geodia megastrella
| Kingston JF & Benson E & Gregory B & Fallis AG 1979: 528 |
Geodia normani, Burton, 1930 : p. 490
| Burton M 1930: 490 |
Sidonops macandrewii
| Brondsted HV 1932: 5 |
| von Lendenfeld R 1903: 100 |
| Sollas WJ 1889: 277 |
normani, Sollas, 1888 : p. 263
| Vosmaer GCJ 1933: 102 |
| Norman AM 1893: 347 |
| Sollas WJ 1888: 263 |
Synops macandrewii
| Sollas WJ 1888: 265 |
Geodia macandrewii, Bowerbank, 1858 : p. 284
| Schottner S & Hoffmann F & Cardenas P & Rapp HT & Boetius A & Ramette A 2013: 2 |
| Murillo FJ & Munoz PD & Cristobo J & Rios P & Gonzalez C & Kenchington E & Serrano A 2012: 842 |
| Cardenas P & Rapp HT & Schander C & Tendal OS 2010: 91 |
| Klitgaard AB & Tendal OS 2004: 57 |
| Klitgaard AB 1995: 2 |
| Cedhagen T 1994: 67 |
| Ereskovsky AV 1993: 23 |
| Barthel D & Tendal OS & Witte U 1991: 40 |
| Hougaard L & Anthoni U & Christophersen C & Larsen C & Nielsen PH 1991: 225 |
| Waren A & Klitgaard A 1991: 52 |
| Koltun VM 1966: 50 |
| Schmidt O 1866: 11 |
| Bowerbank JS 1858: 284 |