
Thelonema clarki, Leduc, 2015
|
publication ID |
https://doi.org/ 10.5852/ejt.2015.158 |
|
publication LSID |
lsid:zoobank.org:pub:16E64AF8-518C-47F0-B3CF-6BA707C222FA |
|
DOI |
https://doi.org/10.5281/zenodo.3793566 |
|
persistent identifier |
https://treatment.plazi.org/id/F4ED998A-656F-48D1-8FBC-B0031BA943D9 |
|
taxon LSID |
lsid:zoobank.org:act:F4ED998A-656F-48D1-8FBC-B0031BA943D9 |
|
treatment provided by |
Carolina |
|
scientific name |
Thelonema clarki |
| status |
sp. nov. |
Thelonema clarki sp. nov.
urn:lsid:zoobank.org:act:F4ED998A-656F-48D1-8FBC-B0031BA943D9
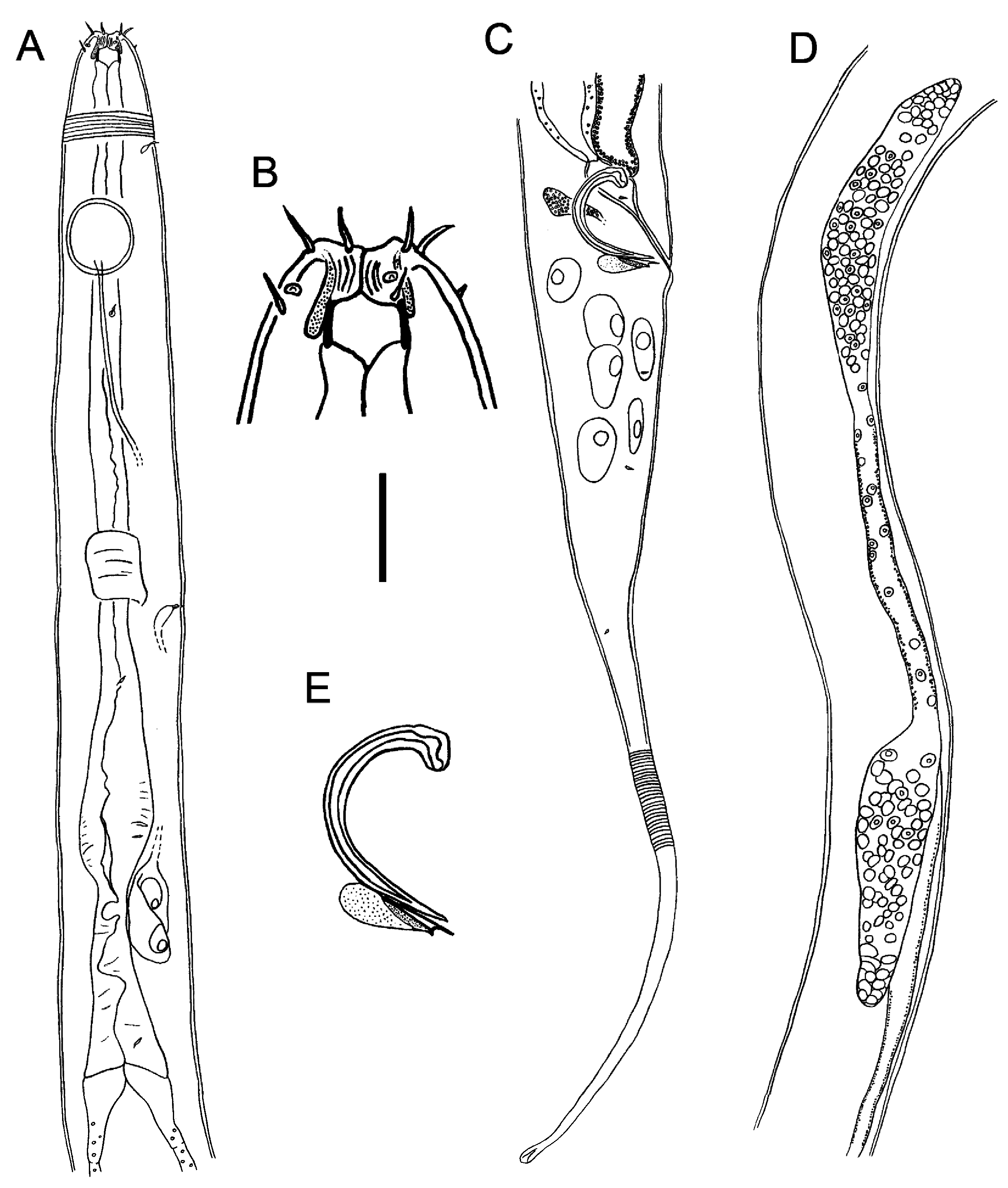
Figs 1–3 View Fig View Fig View Fig , Table 1 View Table 1
Diagnosis
Thelonema clarki sp. nov. is characterised by large adult body size (3230–4461 μm), short cylindrical buccal cavity with cuticularised walls, sub-cephalic setae of similar length or shorter than outer labial setae, secretory-excretory system with two or three renette cells, gubernaculum without apophyses, and conico-cylindrical tail 6.4–10.4 abd long.
Etymology
This species is named after Malcolm R. Clark, principal investigator of the HADES project (HADal Ecosystem Studies) who has made an outstanding contribution to the field of deep-sea ecology and who kindly supported the involvement of the author on the Kermadec Trench voyage.
Material examined
Holotype
KERMADEC TRENCH: ♂ ( NIWA 99760 View Materials ), collected 6 May 2014 (WHOI cruise TN309, Nereus dive N074).
Paratypes
KERMADEC TRENCH: 2 ♀♀ ( NIWA 99761) and 1 juvenile ( NIWA 99762), collected 7 May 2014 ( WHOI cruise TN309, Nereus dive N075).
Type habitat
KERMADEC TRENCH: water depth: 8081 m (178.17571º W, 34.34030º S), sediment depth: 2–3 cm (holotype); water depth: 9177 m (177.65414º W, 32.85037º S), sediment depth: 0–2 cm (paratype).
Description
Male
Body cylindrical, tapering slightly towards both extremities. Cuticle striated from level of buccal cavity to near tail tip. Somatic setae absent except for a few 1 μm long setae in pharyngeal region. Lip region slightly concave; head region otherwise rounded. Internal labial sensillae not observed. Six outer labial setae, 5 μm long, situated in separate circle from the four sensillae of third circle; the latter consists of papillae located in depressions ( Fig. 3B View Fig ). Small, granular glands sometimes observed, apparently connected to base of outer labial setae ( Fig. 1B View Fig ); larger unicellular glands also sometimes observed and apparently connected to cephalic sensillae ( Fig. 2C View Fig ). Four sub-cephalic setae located immediately posterior to the cephalic papillae, 3–4 μm long, similar in length or slightly shorter than outer labial setae. Large circular amphideal fovea with cuticularised outline situated 1.7 cbd from anterior body extremity. Mouth opening narrow, surrounded by bulge of inner portion of lip region; buccal cavity cylindrical, 7 μm deep, 9 μm wide, with cuticularised walls, not surrounded by pharyngeal tissue. Pharynx narrow, muscular, with oval posterior bulb; pharyngeal glands and their orifices indistinct. Nerve ring slightly posterior to middle of pharynx length. Secretory-excretory system with two or three renette cells (third cell may sometimes be obscured) all situated just posterior to pharyngeal bulb; ampulla and pore between nerve ring and posterior pharyngeal bulb. Cardia conspicuous, 90 μm long, widening posteriorly, not surrounded by intestine walls.
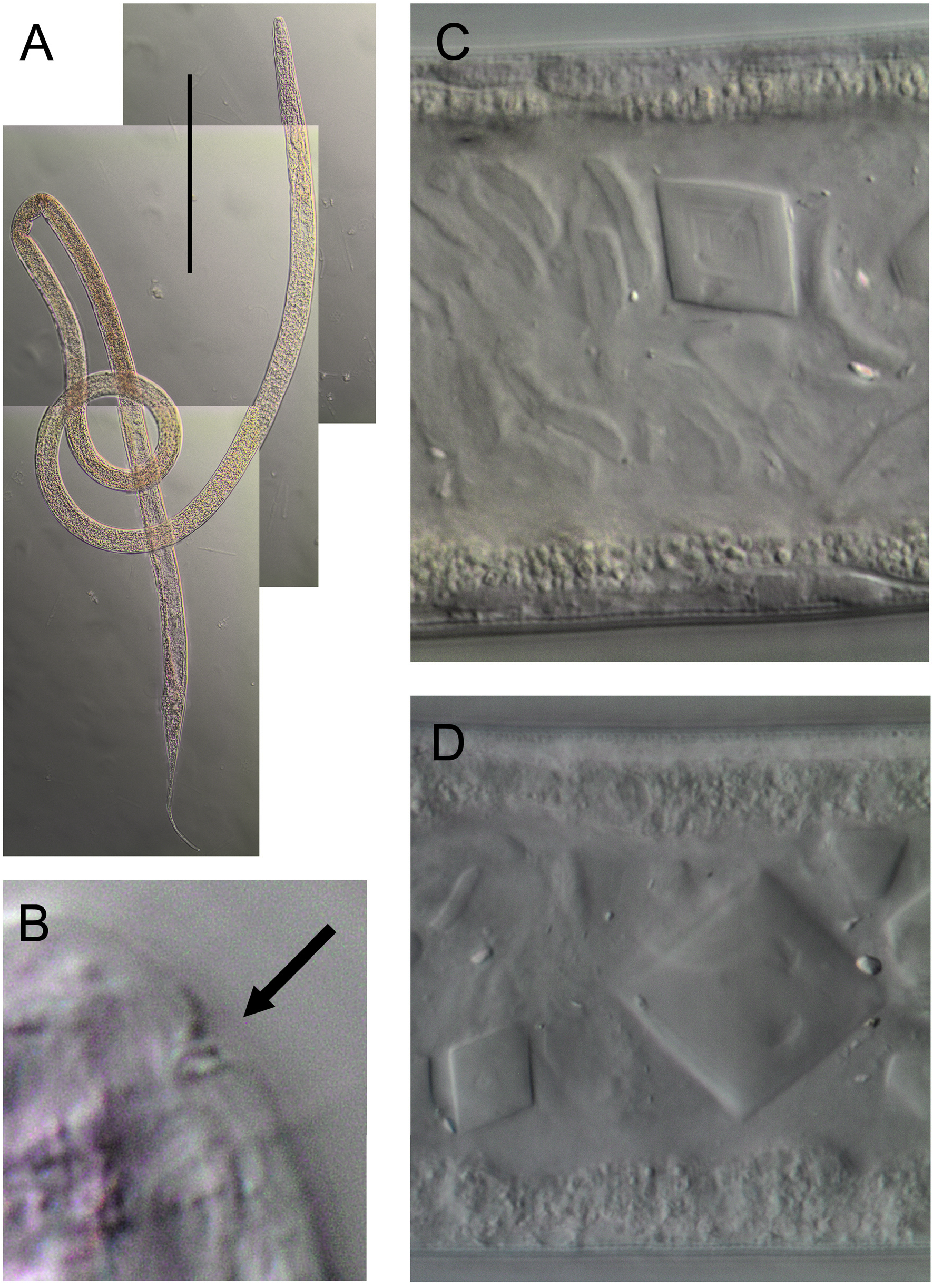
Reproductive system diorchic with two opposed and outstretched testes, anterior testis to the left of intestine and posterior testis to the right of intestine. Mature sperm globular, nucleated, 4–5 × 6–7 μm. Spicules paired, 1.2 abd long, strongly arcuate, with well-developed capitulum and pointed distal end. Gubernaculum thin, pointed at both ends, without apophyses but with rounded glandular tissue extending dorsally. Rectal glands not observed; one ejaculatory gland situated between spicules. Pre-cloacal supplements absent. Tail long, conico-cylindrical, with rounded tip. Caudal glands not observed; short and sparse caudal setae present, no terminal setae. Intestine with numerous transparent crystalline structures, square to rhomboid-shaped, up to 14 × 14 μm ( Fig. 3 View Fig C–D).
Female
Similar to males but with lower values of a, b, and c, slightly smaller amphids (0.42–0.44 vs 0.65 cbd) and longer tail (9.0–10.4 vs 6.4 abd). Buccal cavity 5 μm wide and 4–5 μm deep. Reproductive system didelphic; anterior branch outstretched, to the right of intestine, posteror branch poorly developed, to the left of intestine. Spermatheca present in anterior branch only, simple, not cuticularised. Vulva transverse, situated slightly posterior to mid-body; small vaginal glands present on either side of vagina; muscular pars proximalis vaginae.
Juvenile
Similar to females, but with shorter body and lower values of a and b.
Remarks
Thelonema clarki sp. nov. can be differentiated from the only other species of the genus, T. majum , by the larger body size (3230–4461 vs 1000–1460 μm), short cylindrical buccal cavity (vs long and funnelshaped in T. majum ), sub-cephalic setae of similar length or shorter than outer labial setae (sub-cephalic setae longer than outer labial setae in T. majum ), secretory-excretory system with two or three renette cells (secretory-excretory system not observed in T. majum ), absence of gubernacular apophyses (vs dorso-caudal apophyses present in T. majum ), and longer tail (6.4–10.4 vs 2.7 abd).
This is the first time that Thelonema is recorded outside the type locality in the Peru Basin (~ 4150 m depth) since the original description of the genus by Bussau (1993). The genus was not included in
the recent overview of all Monhysterida by Fonseca & Bezerra (2014), but the discovery of a new Thelonema species in the Southwest Pacific confirms the validity of the genus.
The presence of crystalline structures in the intestine is intriguing, as many appear too large to have been ingested (the structures are up to 14 μm wide, and the width of the cuticularised buccal cavity is 4–9 μm). It seems unlikely that the buccal cavity can stretch to accommodate such large particles given the presence of cuticularised walls. The crystalline structures may have grown through accretion whilst in the intestine, but this process would presumably require a relatively long period of time.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |