
Salmoneus ikaros, Anker & Al-Kandari & Grave, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4780.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:B247849D-2204-436B-A3FC-2E00F0EFF208 |
|
DOI |
https://doi.org/10.5281/zenodo.3853019 |
|
persistent identifier |
https://treatment.plazi.org/id/03F31D2B-FFCA-BC37-FF68-F915FBC7FA2E |
|
treatment provided by |
Plazi |
|
scientific name |
Salmoneus ikaros |
| status |
sp. nov. |
Salmoneus ikaros View in CoL sp. nov.
Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
Material examined. Kuwait: holotype, ovigerous specimen (cl 4.2 mm), OUMNH. ZC. 2020.01.045, Failaka Island , eastern side, 29°23.409’N 48° 23.927’E, mixed rocky-muddy intertidal, muddy patches between rocks, depth less than 0.3 m, suction (yabby) pump, in burrow of ghost shrimp, Balsscallichirus masoomi , leg. A. Anker, 3 October 2019 [fcn KUW-068] GoogleMaps .
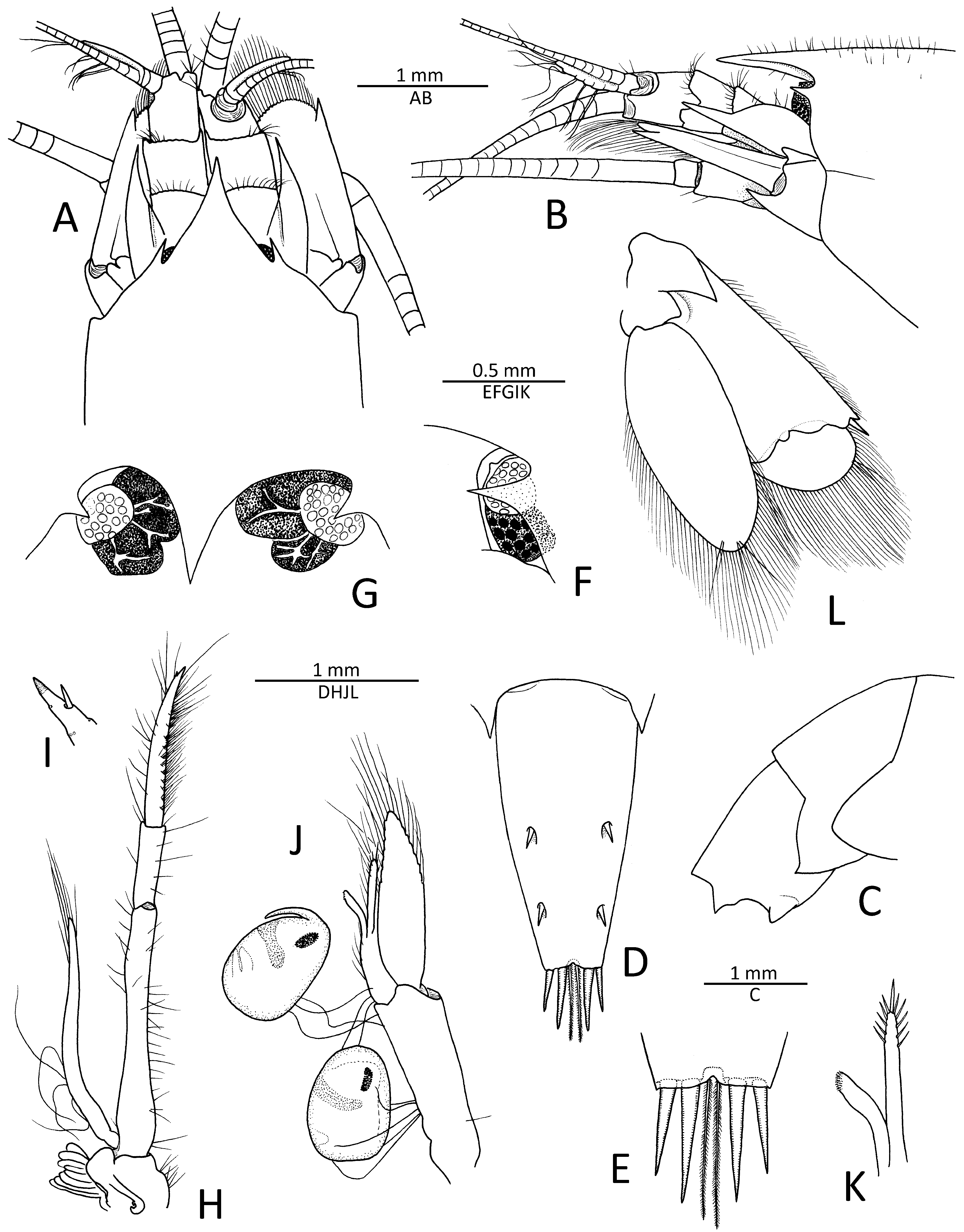
Description.Small-sized(cl 4.2mm)alpheid shrimp with not especially slender or compressed body.Carapace covered with fine erect setae( Fig.1A, B View FIGURE 1 ). Rostrum well developed, broadly subtriangular, slightly longer than wide at base, acute distally, slightly overreaching mid-length of second article of antennular peduncle; lateral margins feebly concave proximally; neither rostral nor mid-dorsal carina present ( Fig. 1A, B View FIGURE 1 ). Orbital teeth well developed, sharp distally, directed slightly mesially in dorsal view, anteriorly in lateral view ( Fig. 1A, B View FIGURE 1 ). Pterygostomial region broadly rounded; anterolateral suture present ( Fig. 1B View FIGURE 1 ); cardiac notch well developed. Each epistomial sclerite with stout acute process.
Pleon with first to fourth pleura broadly rounded to slightly angular; fifth pleuron with posteroventral margin produced into acute, posteriorly directed tooth; sixth pleonite without articulated flap, with short, partly internal suture, posterior margin produced into short subacute tooth flanking telson ( Fig. 1C View FIGURE 1 ); preanal plate posteriorly rounded. Telson moderately slender, subrectangular, tapering distally, almost 2.5 times as long as maximal (proximal) width; dorsal surface with two pairs of small cuspidate setae both inserted at some distance from lateral margin, first pair at about telson mid-length, second pair at about 0.8 of telson length; posterior margin with small shallow notch flanked by with two long plumose setae, remaining margin from central notch to posterolateral angle straight, with two pairs of robust spiniform setae, mesial ones much stouter and about 1.5 times longer than lateral ones ( Fig. 1D, E View FIGURE 1 ).
Eyes almost completely concealed in dorsal view, most-anterior portion exposed between orbital teeth and most basal part of rostrum, partly visible in lateral view; cornea peculiarly divided into two differently pigmented areas, red-brown and black, with branching connections between them ( Figs. 1A, B, F, G View FIGURE 1 , 4 View FIGURE 4 A–C); anteromesial margin with small tubercle ( Fig. 1F View FIGURE 1 ).
Antennular peduncle very stout; stylocerite elongate, with slender distal part, sharp tip almost reaching distal margin of second article; ventromesial carina with small anteriorly directed tooth (not illustrated); second article quadrate, slightly wider than long in dorsal view; lateral antennular flagellum with short fused portion, consisting of three subdivisions, and well-developed secondary ramus, latter with multiple (five) groups of long aesthetascs; mesial antennular flagellum much stouter than lateral ( Fig. 1A, B View FIGURE 1 ). Antenna with basicerite fairly stout, distoventral margin armed with large acute tooth; scaphocerite oval-shaped, not reaching end of antennular peduncle, with straight lateral margin and broad blade, latter convex anteriorly, reaching slightly beyond stout distolateral tooth; carpocerite short, cylindrical, reaching to about mid-length of scaphocerite ( Fig. 1A, B View FIGURE 1 ).
Mouthparts not dissected, appearing typical for genus in external observation. Third maxilliped slender, pediform; coxa with rounded lateral plate; antepenultimate article slender, slightly flattened ventrolaterally; penultimate article about four times as long as wide; ultimate article with numerous rows of short serrulate setae and some longer simple setae, tip with subacute corneous point and one subapical spiniform seta; arthrobranch modestly developed ( Fig. 1H, I View FIGURE 1 ).
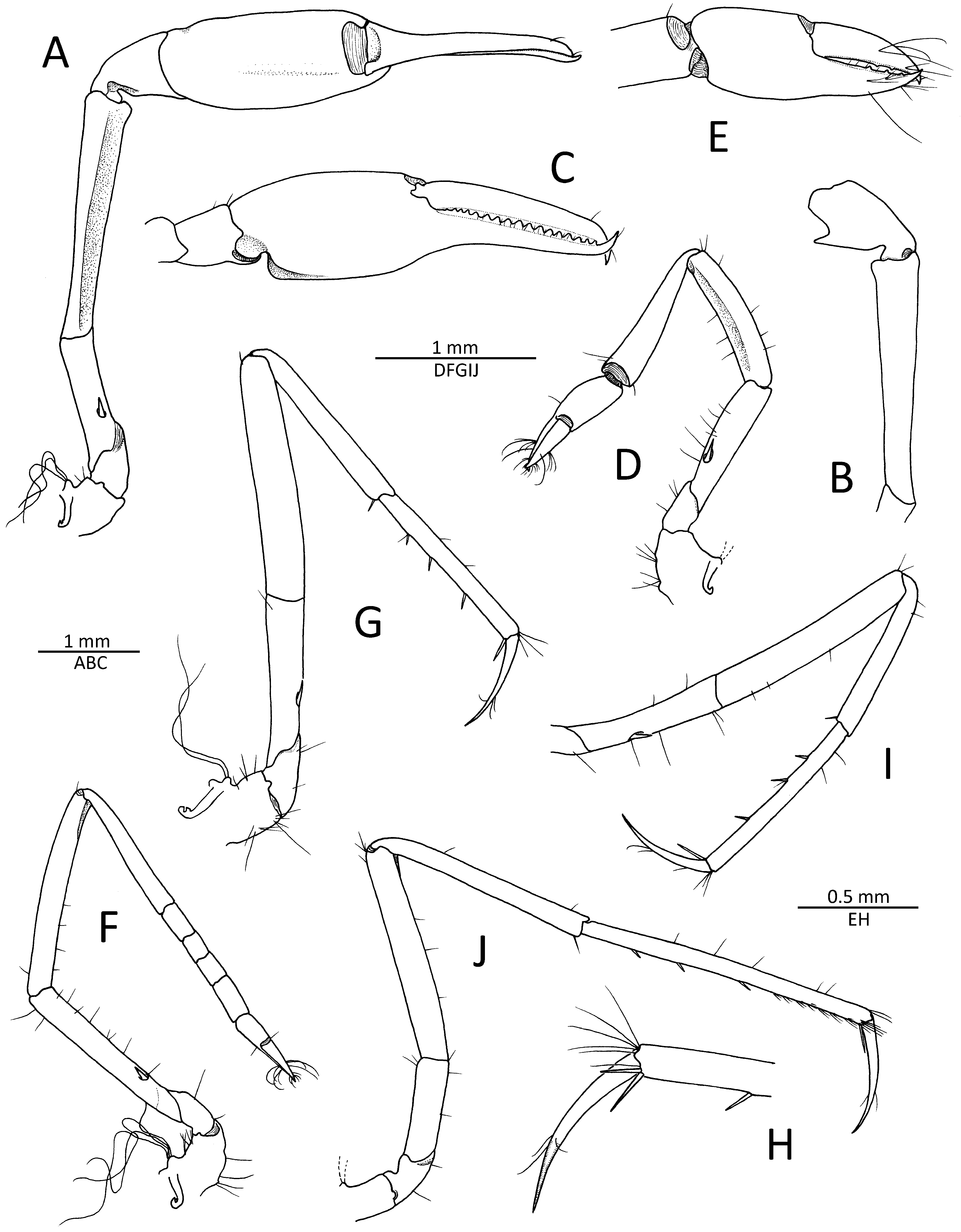
First pereiopods (chelipeds) very asymmetrical in shape and dissimilar in size, carried flexed when not in use ( Figs. 2 View FIGURE 2 A–E, 3A, B). Major cheliped enlarged, with slender ischium and merus and moderately swollen chela; ischium flattened ventrolaterally, armed with one cuspidate seta; merus more than seven times as long as proximal width, somewhat broader distally, smooth, distodorsal and distomesial margins rounded, ventrolateral surface depressed to accommodate chela when flexed; carpus short, cup-shaped, with distal margin bluntly protruding towards propodus of chela mesioventrally, but without sharp teeth or processes; chela fairly large relative to body, as long as all proximal articles combined; palm subcylindrical smooth, without grooves or sculpture, except for deep proximal constriction on ventral surface; fingers slightly longer than palm; not gaping when closed, subequal in length, slightly twisted and crossing distally, with evenly serrated cutting edges; both dactylus and pollex with about 17 rounded to subtriangular teeth on cutting edge, extending for most of finger length, except for distal-most portion ( Fig. 2 View FIGURE 2 A–C). Minor cheliped substantially smaller than major cheliped, less robust; ischium armed with one cuspidate seta on ventrolateral surface; merus subequal to ischium, not swollen or widening distally, smooth, slightly depressed ventrally; carpus about 1.2 times as long as merus, cylindrical, widening distally; chela much shorter than carpus, simple, with palm about as long as fingers, smooth; fingers somewhat gaping when closed, subequal in length, crossing distally, cutting edges with three minute, widely spaced teeth on distal half ( Fig. 2D, E View FIGURE 2 ).
Second pereiopod slender; ischium with one cuspidate seta on ventrolateral surface, situated in proximal third; merus longer than ischium; carpus with five subdivisions, first about as long as four others combined, their ratio approximately equal to 4.0/1.0/0.7/0.8/1.3; chela longer than distal-most carpal subdivision, simple ( Fig. 2F View FIGURE 2 ). Third pereiopod moderately slender; ischium with one cuspidate seta on ventrolateral surface; merus about six times as long as wide, unarmed; carpus noticeably slenderer and shorter (at most 0.7 times) than merus, with one slender spiniform seta on distoventral margin; propodus longer than carpus, with three widely spaced spiniform setae on ven- tral margin, in addition to one pair of somewhat longer spiniform setae (ventrolateral much longer than ventromesial) near dactylar base; dactylus overreaching half-length of propodus, slender, conical, simple, smoothly curving distally ( Fig. 2G, H View FIGURE 2 ). Fourth pereiopod similar to third pereiopod in armature and article length proportions, slightly more slender ( Fig. 2I View FIGURE 2 ). Fifth pereiopod as slender as fourth pereiopod; ischium unarmed; merus at least seven times as long as wide, unarmed; carpus more slender than merus, about 0.9 length of merus, with stiff seta distoventrally; propodus very long, slender, 1.3 times as long as carpus, with eight or so rows of progressively longer serrulate setae forming cleaning brush on distal ventrolateral surface, ventral margin with three spiniform setae, in addition to one pair of longer spiniform setae near dactylar base (mesial invisible in lateral view); dactylus about 0.4 times length of propodus, similar to that of third and fourth pereiopods ( Fig. 2J View FIGURE 2 ).
Second pleopod with appendix masculina almost twice as long as appendix interna, furnished with numerous long stiff setae, apical stoutest, as illustrated ( Fig. 1J, K View FIGURE 1 ). Uropod with lateral lobe of protopod ending distally as large subacute tooth; exopod ovate, with small subacute distolateral tooth and well-developed distolateral spiniform seta; diaeresis sinuous, with broadly rounded tooth mesial to distolateral spiniform seta; endopod as long as exopod, narrowly ovoid, without specific features ( Fig. 1L View FIGURE 1 ).
Gill formula typical for genus.
Colouration. Semi-transparent, white, with dull reddish and black cornea and yolk-yellow eggs ( Fig. 3 View FIGURE 3 ).
Etymology. The new species is named after the ancient Greek name of the type locality, Ikaros, currently known as Failaka Island, Kuwait; used as a noun in apposition.
Distribution. Indian Ocean: presently known only from the type locality in Kuwait.
Ecology. The holotype, and single known specimen, was extracted from a burrow of the ghost shrimp Balsscallichirus masoomi ( Tirmizi, 1970) (Callianassidae) , together with its host (OUMNH.ZC. 2020.01.036 [fcn KUW- 067], material to be reported in more details elsewhere), on a shallow subtidal flat characterised by murky water and fine sand-mud bottom, near the edge of a large rocky outcrop ( Fig. 9 View FIGURE 9 ).
Remarks. Salmoneus ikaros sp. nov. differs from all other species described in the genus Salmoneus by its highly peculiar configuration of the eyes ( Figs. 1F, G View FIGURE 1 , 4 View FIGURE 4 A–C). The dendrite-like connections between the two differently pigmented areas are especially noteworthy. It is possible that the oddly shaped eyes of S. ikaros sp. nov. represent an adaptation to the light-deficient microhabitat of the species, i.e. deep galleries of ghost shrimps, in addition to fairly murky waters due to the mud and silt-dominated bottom. The only other species of Salmoneus that has somewhat atypical eyes is S. komaii Anker, 2011 , in which the cornea possesses an unusual lateral lobe ( Anker 2011: fig. 5E). However, S. komaii has very differently shaped chelipeds and is only remotely related to S. ikaros sp. nov.
Anker & Marin (2006) established several informal species groups within Salmoneus , mainly based on the features of the rostro-orbital region and chelipeds. However, the dramatic increase of both number of species and morphological heterogeneity within the genus made at least some of these groupings artificial and/or obsolete, and it is now clear that some of them are non-monophyletic. Many species described since 2006 presented characters not in line with the species group definitions provided by Anker & Marin (2006), whereas others present intermediate or novel features (e.g., Anker 2011; Komai & Anker 2012; Anker et al. 2014; Ďuriš & Horká 2016). Nevertheless, these informal groups still have utility as a first approximation, i.e. initial assessment of the morphological characters of a given species and its possible relationships.
Salmoneus ikaros sp. nov. appears to be morphologically closest to the species of the S. gracilipes Miya, 1972 species group, as defined by Anker & Marin (2006), based on the dorsally partly visible eyes and the general shape of the chelipeds. This group originally included, in addition to S. gracilipes , the following species: S. colinorum De Grave, 2004 , S. alpheophilus Anker & Marin, 2006 , S. pusillus Anker & Marin, 2006 , S. falcidactylus Anker & Marin, 2006 , S. seticheles Anker, 2003 , S. cavicolus Felder & Manning, 1986 , and possibly S. tafaongae Banner & Banner, 1966 ( Anker & Marin 2006). Among the species described since 2006, only S. hispaniolensis Anker, 2010 , S. camaroncito Anker, 2010 and S. antricola Komai, Yamada & Yunokawa, 2015 could be assigned to the S. gracilipes group, based on their overall morphology ( Anker 2010; Komai et al. 2015), although several characters of the latter two species do not totally fit into the definition of this group.
Among the above-mentioned species, S. gracilipes , S. colinorum , S. alpheophilus , S. pusillus , S. falcidactylus , S. seticheles , S. tafaongae and S. antricola occur in the Indo-West Pacific. Asides from its unique eyes, S. ikaros sp. nov. can be distinguished from all of them by the shorter and typically proximally broader rostrum, as well as a number of other characteristics, e.g., from S. gracilipes by the absence of a longitudinal depression on the major chela, the absence of a rostral carina and the presence of only one spiniform seta on the ischia of the third and fourth pereiopods (vs. three in S. gracilipes ) (cf. Miya 1972); from S. colinorum , S. alpheophilus , S. falcidactylus and S. seticheles by the longer stylocerite, almost reaching the distal margin of the second article of the antennular peduncle (vs. reaching or slightly overreaching its mid-length in the other species) (cf. Anker 2003; De Grave 2004; Anker & Marin 2006); from S. colinorum by the uniform whitish colour of the body (vs. with purple-brown transverse bands on the carapace and pleon in S. colinorum ) (cf. De Grave 2004; Anker et al. 2015; Anker 2019); from S. alpheophilus by the absence of a post-rostral tubercle, and the much longer appendix masculina, almost twice as long as the appendix interna (vs. shorter than the appendix interna in S. alpheophilus ) (cf. Anker & Marin 2006); from S. pusillus by the absence of a subdistal tooth on the ventral margin of the rostrum and post-rostral tubercle (vs. present in S. pusillus ), the presence of a small notch on the posterior margin of the telson (vs. absent in S. pusillus ), and the stouter walking legs (third to fifth pereiopods), including the much shorter carpus of the third pereiopod (at most 0.7 as long as merus vs. as long as merus in S. pusillus ) (cf. Anker & Marin 2006); from S. falcidactylus by the ischia of the chelipeds and second pereiopods armed with cuspidate setae (vs. unarmed in S. falcidactylus ), the much more robust major chela and the stouter walking legs, including the shorter, more robust dactyli (vs. extremely slender, sickle-shaped dactyli in S. falcidactylus ) (cf. Anker & Marin 2006); from S. seticheles by the absence of long fringing setae on the major chela (vs. present in S. seticheles ) and the presence of armature on the major and minor cheliped ischia (vs. absent in S. seticheles ) (cf. Anker 2003); from S. tafaongae by the anteriorly (not upwards) directed orbital teeth (cf. Banner & Banner 1966); and from the highly derived, cavernicolous S. antricola by the much more robust major cheliped, with a very different serration on the cutting edges of the fingers, and several other characters (cf. Komai et al. 2015).
Salmoneus ikaros sp. nov. can also be separated from all other Indo-West Pacific species of Salmoneus , including all members of the S. serratidigitus (Coutière, 1897) and S. cristatus (Coutière, 1897) groups, which highly likely also represent non-monophyletic groupings in their present compositions. The species morphologically closest to S. ikaros sp. nov. may be the poorly known S. latirostris (Coutière, 1897) , which has a very similar rostroorbital region of the carapace, including the anterior-most portion of the eyestalk being exposed dorsally ( Coutière 1899: fig. 21). However, S. latirostris has a faint rostral carina and is red-banded in life ( Coutière 1899), unlike our new species from Kuwait. The recently described S. venustus Anker, 2019 , also a symbiont of callianassid burrows, differs from S. ikaros sp. nov. by the much shorter stylocerite, the presence of conspicuous anteromesial tubercles on the eyestalks, the more produced lateral plate of the third maxilliped, the absence of ischial armature on the chelipeds and all walking legs, as well as by the red-banded colour pattern (cf. Anker 2019).



| ZC |
Zoological Collection, University of Vienna |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |