
Pararhynchelmis murdocki, Fend, Steven V. & Lenat, David R., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.196882 |
|
DOI |
https://doi.org/10.5281/zenodo.6206487 |
|
persistent identifier |
https://treatment.plazi.org/id/03EEF043-2A3B-6B7A-A1D3-FA0BFE92A340 |
|
treatment provided by |
Plazi |
|
scientific name |
Pararhynchelmis murdocki |
| status |
sp. nov. |
Pararhynchelmis murdocki View in CoL n. sp.
( Figures 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 )
Holotype: USNM 1142479, a longitudinally dissected worm, stained in hematoxylin and slide mounted.
Type locality: USA, Virginia, Amherst Co., small spring in Peaks of Otter area, near Blue Ridge Parkway Milepost 85.5, behind maintenance area, elevation 786 m, N37° 27' 21.30" W79° 35' 56.58", 7 May 2007, collected by D.R. Lenat.
Paratypes: The type locality, 7 May 2007: USNM 1142480-1142483, 1 whole mount, 1 dissected, 1 sagittally sectioned, 1 transversely sectioned.
Other material: USA: Virginia: Amherst Co., springs in Peaks of Otter area, Blue Ridge Parkway. The type locality, 7 May 2008: 3 whole mounts, 3 dissected, 1 sagittally sectioned, 1 transversely sectioned. 13 April 2008: 3 whole mounts, 9 dissected. Big Spring and associated seeps, near Milepost 86.5, elevation 752 m, N37° 26' 33.12" W79° 35' 50.82", 7 May 2007: 2 whole mounts, 9 dissected. 14 April 2008: 2 whole mounts, 7 dissected. All collected by D.R. Lenat.
Etymology. Named for Nora Murdock, a National Park Service biologist responsible for conducting biological surveys along the Blue Ridge Parkway of Virginia and North Carolina.
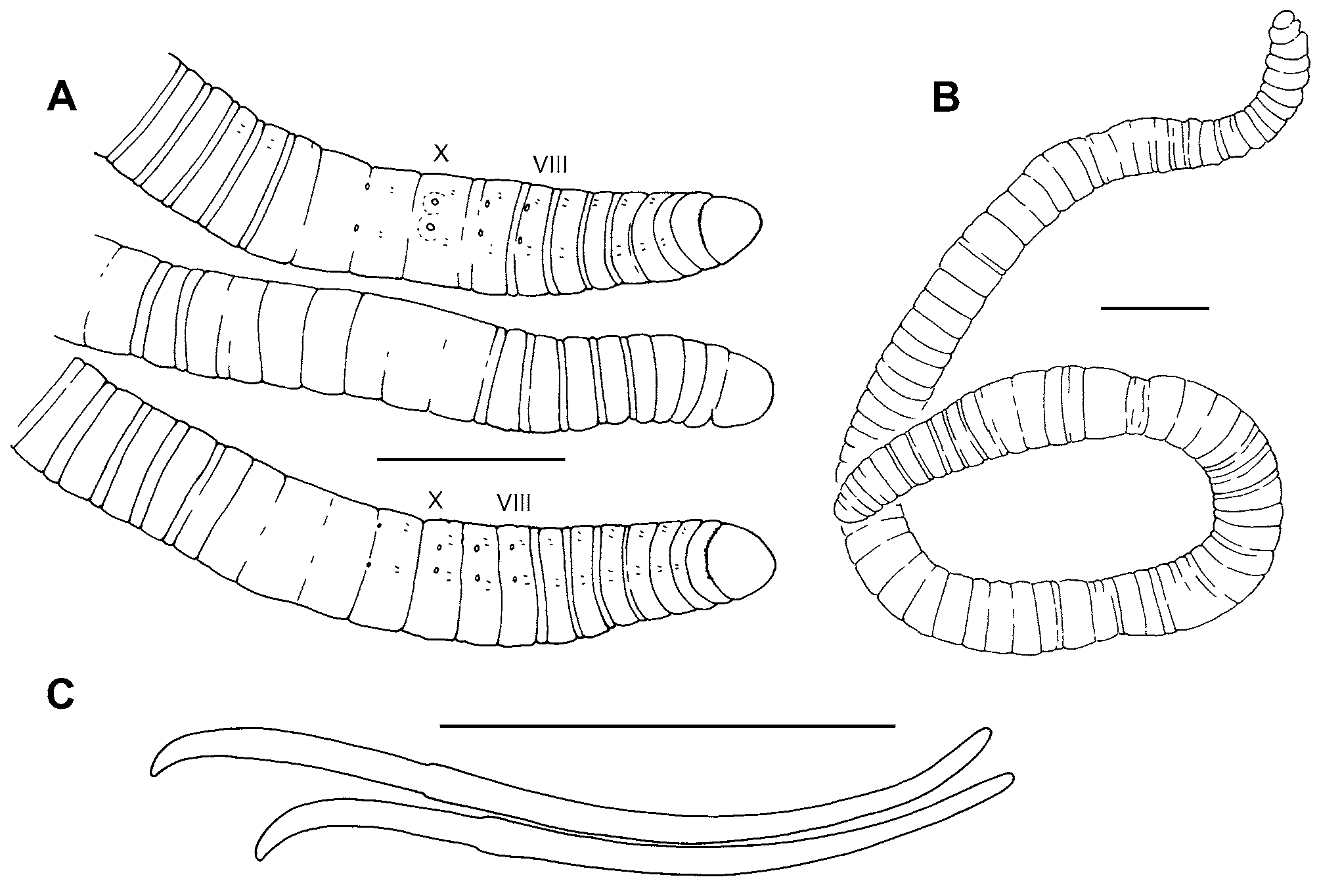
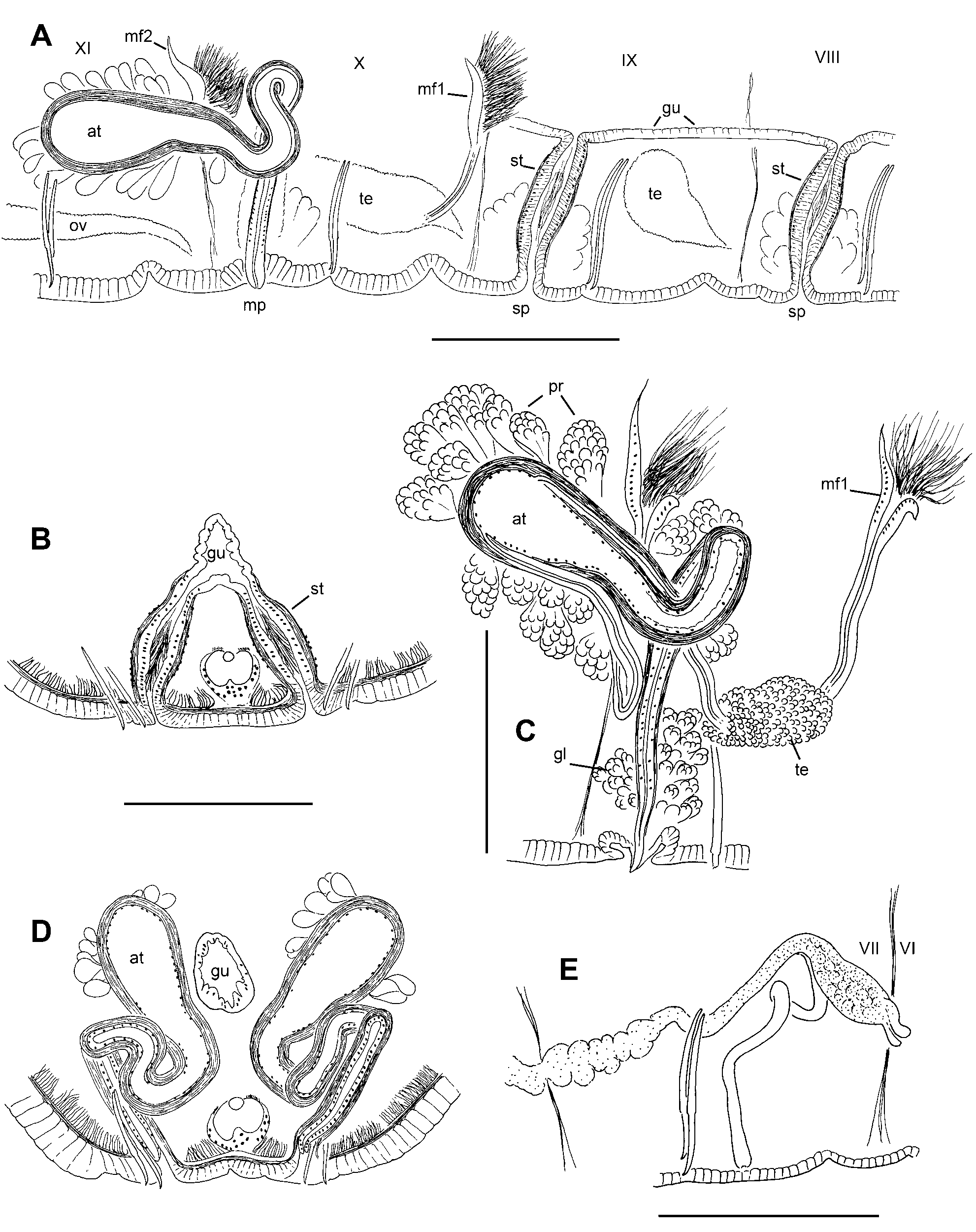
Description. Length of preserved worms 18 (14–25) mm; segments 87 (76–103); diameter 0.50 (0.35– 0.71) mm in X, maximum diameter 0.57 (0.45–0.73) mm ( Fig. 7 View FIGURE 7 A–B). Prostomium rounded, prolobous, slightly wider than long. Secondary segmentation a narrow anterior ring in IV–VIII, annulations may be externally indistinct in clitellum; secondary segmentation variable in post-clitellar segments. Epidermis 20–30 μm thick in prostomium, 7–14 μm in preclitellar area, up to 25–38 μm in clitellum, and 5–10 μm posteriorly. Longitudinal muscle 20–50 μm thick, usually thicker ventrally than dorsally; circular muscle 2–4 μm. Clitellum usually mid-VIII-XIII, thickest in X–XI; not developed ventrally near genital pores. Chaetae simple-pointed, moderately sigmoid; nodulus approximately 1/3 the distance from tip (0.29–0.37) ( Fig. 7 View FIGURE 7 C). Chaetal length 113–170 μm in clitellar region, 115–158 μm posteriorly; dorsal and ventral chaetae approximately the same, or dorsals may be slightly shorter in anterior segments. Pharynx from II–III, dorsal and lateral walls distinctly thickened, midventral area thin ( Fig. 9 View FIGURE 9 A). Pharyngeal glands in dorsolateral and ventrolateral cell clusters near the posterior septa in IV–VI, sometimes extending into VII. Brain in the peristomium.
First nephridia usually paired on 6/7; second pair usually on 12/13; nephridia occur irregularly (approximately every third to fourth segment on either side) in posterior segments. Each nephridium with small anteseptal funnel; an ovate postseptal expansion (length about 60–90 μm, diameter 40 μm); and a long, convoluted duct, which usually loops posteriorly through two or more segments ventral to the gut, and terminates in a nephropore anterior to the ventral chaetae in the originating segment ( Fig. 8 View FIGURE 8 E). Ectal ducts may widen slightly at the nephropore, but do not form a distinct vesicle.
One pair of thin, sinuous commissural blood vessels in II–VI; generally joining ventral vessel in the segment posterior to their origin in the dorsal vessel. Commissures shorter and less convoluted in VII–IX; those in X and XI may loop posteriorly within sperm and egg sacs. Posterior segments from about XX to XL have a single pair of thick, lateral blood vessels near the anterior septum, joining the dorsal vessel to the ventrolateral side of the gut. Posterior to about XL, a second pair of unbranched lateral vessels occurs in the posterior part of each segment. Perivisceral sinus and chloragogen begin in about VII.
Testes paired in IX and X; those in X large, extending to posterior part of segment; those in IX usually slightly smaller. Ovaries in XI; elongate, usually extending through XII. Sperm sacs extend forward as far as VII or VIII, posteriorly to XV–XVI. Egg sacs extend back an additional 1–3 segments when mature eggs are present. Female pores intersegmental on 11/12, in line with ventral chaetae; female funnels simple, cupshaped, about 80–90 μm high. Spermathecae paired in VIII and IX; atria paired in X; both male and spermathecal pores posterior and slightly median to the ventral chaetae ( Fig. 7 View FIGURE 7 A).
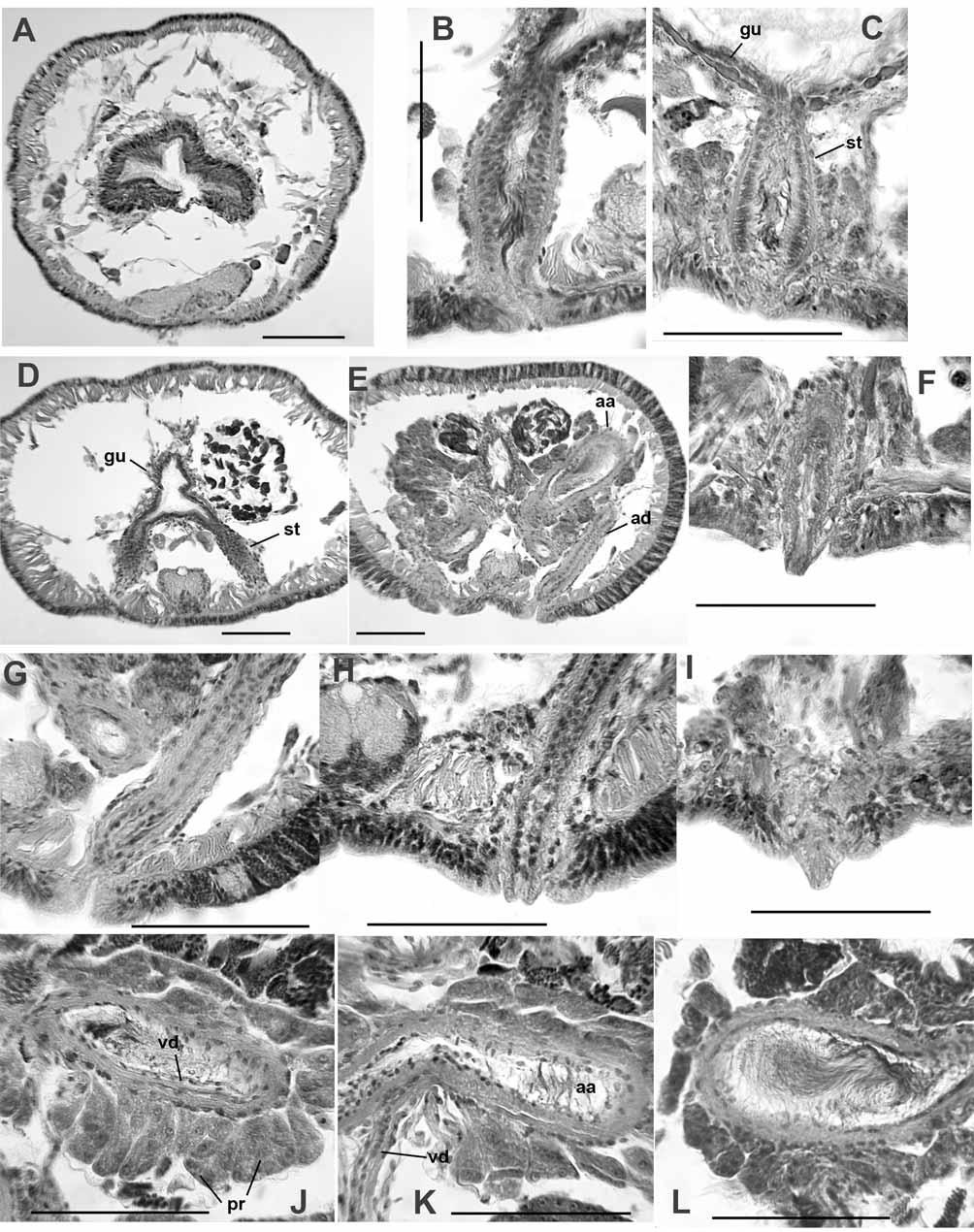
Spermathecae are simple tubes, slightly tapered at ends; structure duct-like, without distinct ampulla or diverticula; ental ends join and open into ventrolateral side of gut ( Fig. 8 View FIGURE 8 A–B, 9B–D). Size and morphology similar in VIII and IX; length 137 (96–205) μm; diameter at middle 40–60 μm. A thin muscle layer (2–4 μm) surrounds the spermatheca; epithelial cells densely packed, somewhat columnar; lumen narrow, to 10 μm ( Fig. 9 View FIGURE 9 B–C). Ectally, the spermatheca terminates in a simple and inconspicuous pore; internally, the pore area with anterior and posterior masses of loosely clustered, petiolate gland cells, either single or in small bundles resembling prostates ( Fig. 9 View FIGURE 9 C). Sperm cells loosely packed, with heads oriented towards epithelium in ectal part of spermatheca Fig. 9 View FIGURE 9 B–C).
Male funnels on 9/10 and 10/11, the anterior pair simple, 67–100 μm high; the posterior pair usually slightly larger, 72–130 μm high, sometimes convoluted or folded back into sperm sacs. In some specimens both pairs of male funnels functional, with similar amount of sperm; in other worms the anterior funnels without sperm or with much less than the posterior pair. Vasa deferentia very thin, diameter 6–8 μm. Posterior vasa do not form a loop in the postatrial segment or penetrate septum 10/11. Both vasa deferentia join the atria near the midpoint (near the base of the ampulla), run under the atrial muscle layer, and enter the atrial lumen subapically ( Fig. 8 View FIGURE 8 C, 9J–K).
Atria elongate-petiolate, indistinctly divided near the midpoint into a tubular, ectal duct (length 190–370 μm, width 26–38 μm) and an ental, club-shaped ampulla (length 180–340 μm; maximum width to 72–115 μm near the ental end)( Fig. 8 View FIGURE 8 C, 9E). The duct rises vertically from the male pore and forms a loop within X ( Fig. 8 View FIGURE 8 C–D). The ampulla almost always exends posteriorly into XI within a sperm sac, and is distinguished from the duct by the slightly thicker muscle layer and presence of prostates. Atrial epithelium in ectal duct 3–5 μm thick, with cuboidal cells; epithelium in the ampulla composed of larger, vacuolated cells having basal nuclei, but cells often appear eroded, and may be indistinct ( Fig. 9 View FIGURE 9 L). Atrial muscle layer made up of mostly transverse-circular fibers, 8–12 μm thick in ampulla; thinner (5–6 μm) in the duct. Prostate glands loosely cover the ampullar portion in distinct, pyriform bundles of cells, 35–60 μm long, usually longest in ectal part of ampulla; prostate cells indistinct and granular ( Fig. 8 View FIGURE 8 C, 9J–L).
Ectal duct of atrium terminates in a short (20–30 μm), conical penis ( Fig. 9 View FIGURE 9 F,I), which may be indistinct when retracted into a shallow sac ( Fig. 9 View FIGURE 9 G). Penes are short extensions of the atrial duct, apparently surrounded by extensions of epidermal cells ( Fig. 9 View FIGURE 9 H). Male pore internally surrounded by a loose cluster of petiolate gland cells ( Fig. 9 View FIGURE 9 F).
Remarks. The location of the spermathecae, paired in the first two preatrial segments, is unique within the Nearctic lumbriculid fauna. Morphology of the short, undifferentiated spermathecae (without distinct ducts, ampullae, or diverticula) is unlike that of any other lumbriculid. Despite similarities in arrangement of the reproductive organs, and the junction of the spermathecae with the gut, P. murdocki differs from typical Rhynchelmis in most respects (see Discussion). It lacks a proboscis, body size is small, lateral blood vessels are unbranched, nephridia are present on septum 6/7, anterior sperm sacs are present, the atria are shorter and more distinctly petiolate, and the anterior male ducts are functional.
Pararhynchelmis murdocki has only been collected in two springs located in a relatively small area near the Peaks of Otter Mountains in Virginia (Lenat 2007). The worms are most abundant where the springs emerge from the ground, but it is not known whether this species is limited to a very restricted habitat at the head of the spring or if they are part of a large groundwater population. Similar springs farther north along the Blue Ridge Parkway (miles 6–29) also contain large numbers of lumbriculids, but do not contain this species. Most worms were fully or partially mature on the few collection dates, from mid-April to early May. As P. murdocki appears restricted to spring habitats, it may require year-round flow and a relatively constant temperature.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |