
Sinobatis kotlyari, Stehmann, Matthias F. W. & Weigmann, Simon, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4189.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:2A278CBF-493D-423C-8ED3-2E7AE362C2E4 |
|
DOI |
https://doi.org/10.5281/zenodo.5691291 |
|
persistent identifier |
https://treatment.plazi.org/id/03E9879A-8337-FFEA-4EF3-1D94FCEBFED6 |
|
treatment provided by |
Plazi |
|
scientific name |
Sinobatis kotlyari |
| status |
sp. nov. |
Sinobatis kotlyari View in CoL n. sp.
(Kotlyar's legskate)
( Figures 2–20 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 ; Table 1)
Sinobatis sp. Vietnam—Weigmann et al., 2014: pp. 123, 124, 130.
Sinobatis sp. Broken Ridge—Weigmann & Stehmann, 2016: pp. 495, 499.
Holotype: ZMMU P-17178, almost adult male 331 mm TL; RV “Professor Mesyatsev”, cr. 7, 1 April 1979, trawl 24 (bottom), Broken Ridge, 30°50’ S, 92°29’ E, 1400 m; collector Dr. A. Kotlyar.
Diagnosis. A small species of the genus Sinobatis distinguished by the cranial feature of having very elongated and extremely narrow fontanelle at its anterior two thirds, but mainly by characters of its clasper. Disc broadly inverse heart-shaped with slightly convex anterior margins and broadly rounded outer and inner pectoral corners; width 64% of TL, length 63% of TL. Head (dorsal length 27% of TL, ventral length 34% of TL) and snout (preorbital length 22% of TL, preoral length 23% of TL, prenasal length 21% of TL) moderately long; snout without rostral filament and angled at 75°. Tail cord-like, hardly tapering towards its tip, and slightly shorter than body. No dorsal fins, caudal fin rather well developed, base lengths of its upper and lower lobes 8% of TL and 6% of TL, respectively. Upper side largely plain medium greyish-brown, underside generally lighter than upper surface, beige with darker outer and posterior areas. Clasper glans with narrow shield, long and distinct pent, slender and smooth-edged distal sentinel and spike but no other cartilage-supported components on outer or inner terminal lobes; clasper skeleton lacking dorsal terminal 1 but with elongated dorsal terminals 2 and 3, narrow twisted ventral terminal with long and curved proximal process, and long, slender, pointed accessory terminals 1 and 2 inserting side by side at inner distal edge of ventral marginal. The combination of colouration, several morphometrics and meristics, as well as few external clasper components and terminal skeletal cartilages distinguishes the species from congeners.
Description of holotype. Detailed morphometrics and meristics given in Table 1. Where relevant, ratios are based on horizontal measurements unless otherwise stated.
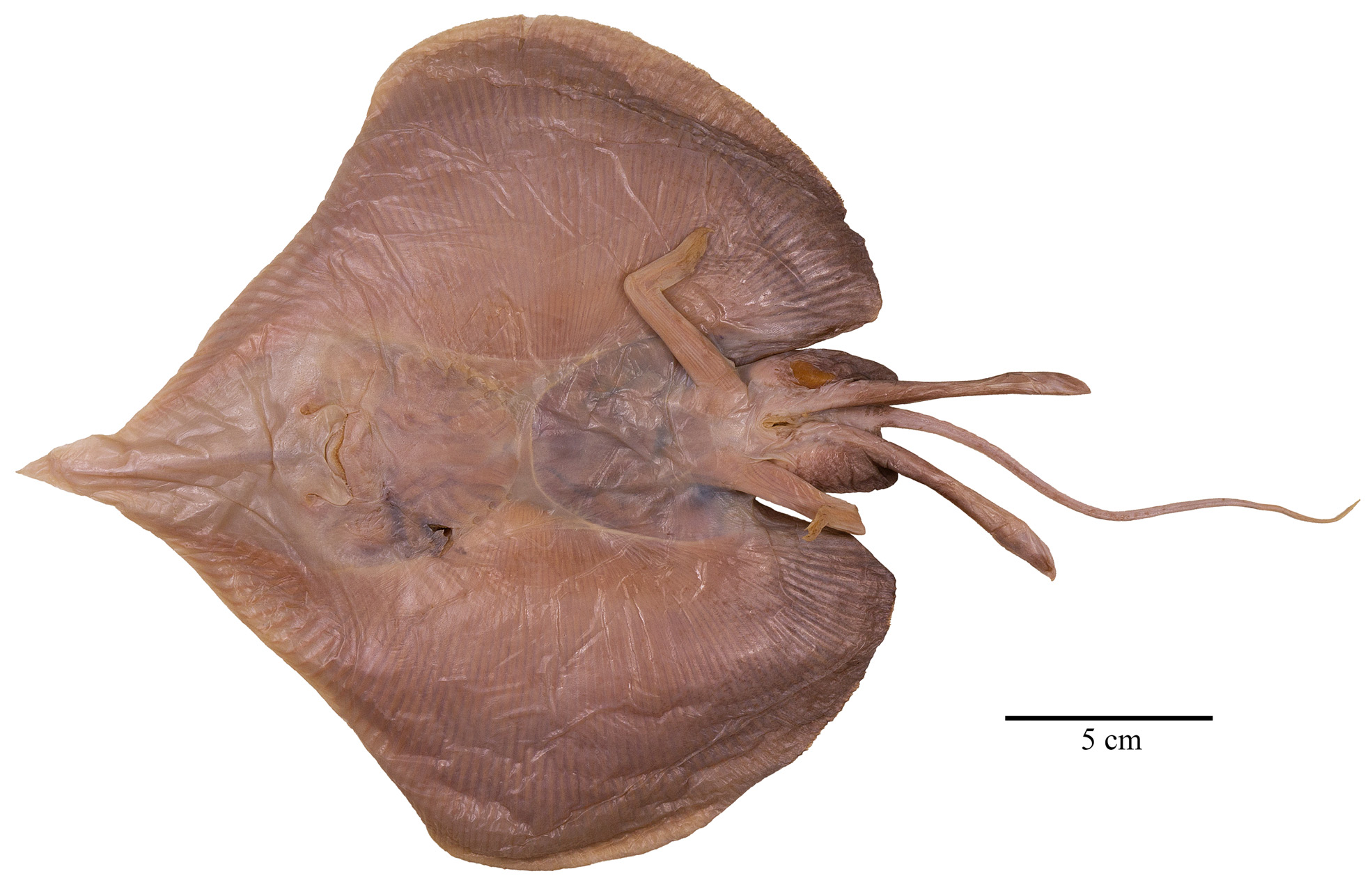
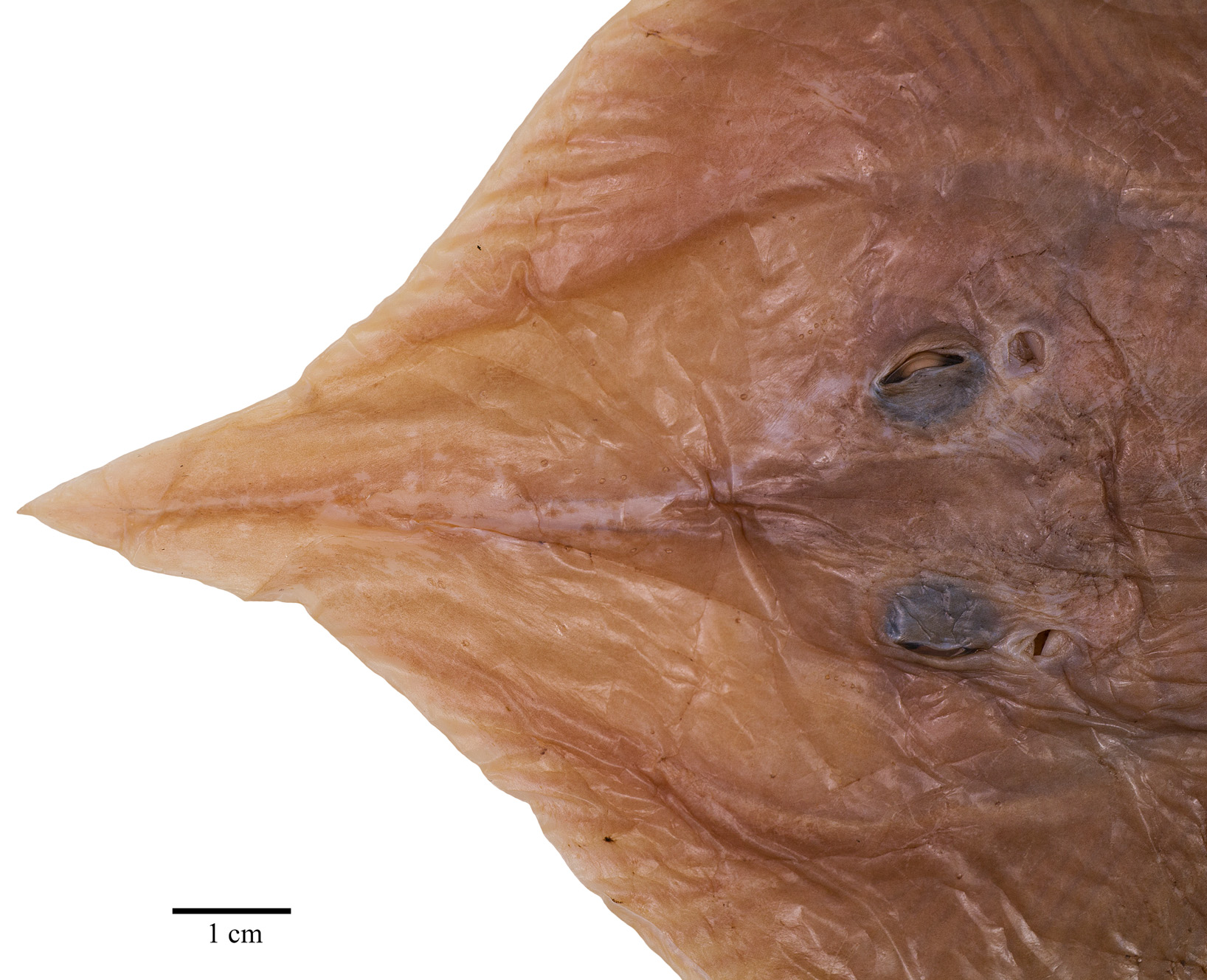
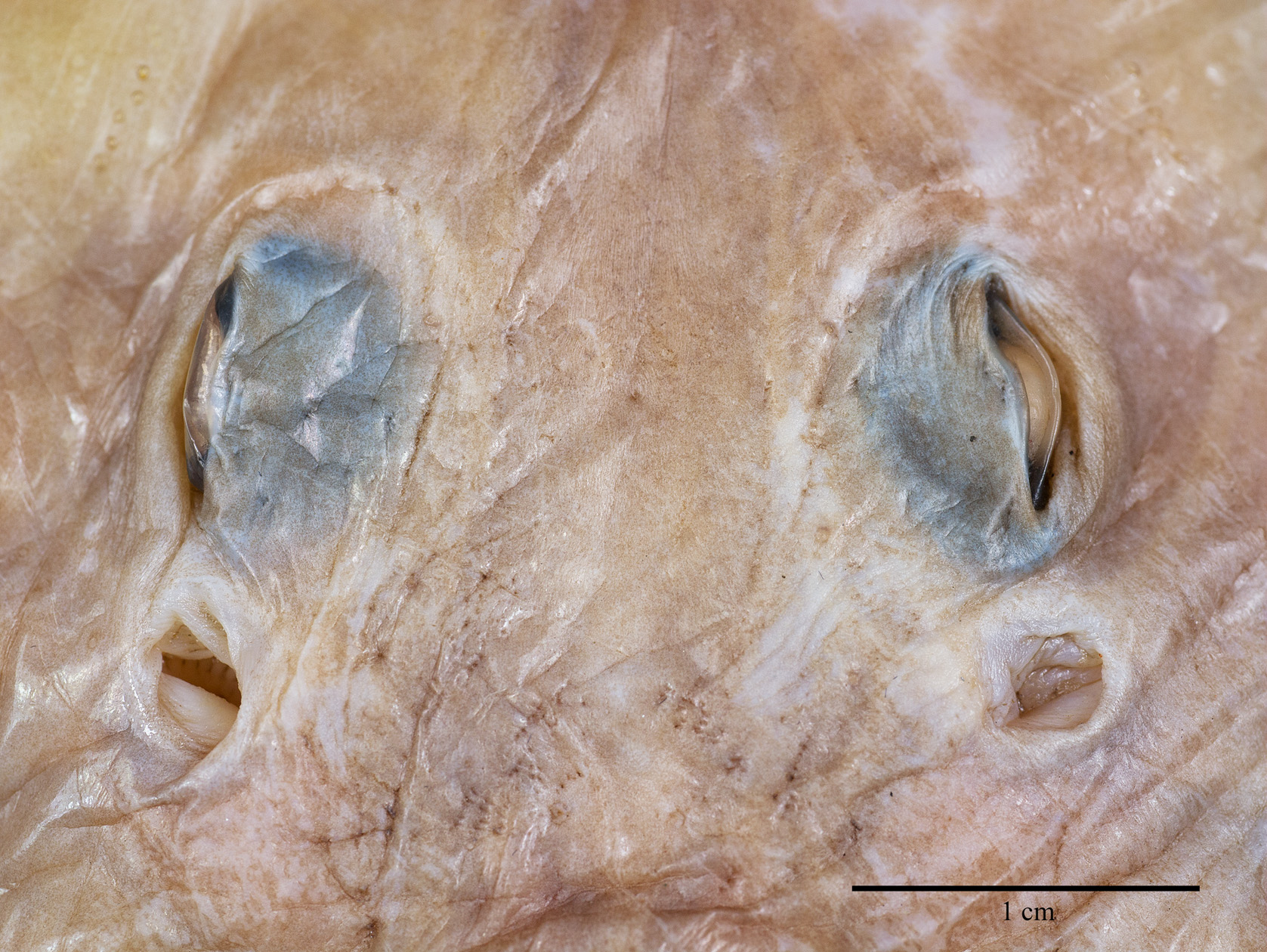
External morphology ( Figs. 2–11 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 ). Disc broadly inverse heart-shaped; anterior margins slightly convex, outer and inner pectoral corners broadly rounded, posterior margins evenly convex, pectoral axils not fused to outer margin of posterior pelvic lobes; disc width almost equal to disc length, axis of maximal disc width at 35% of TL and 55.3% of disc length. Dorsal head length 27% of TL and 43.7% of disc length. Snout long and pointed, gradually tapering to the rostral lobe which is not marked-off; rostral filament absent; snout narrowly pointed at about 75°; preorbital snout length 2.9 times in disc length and 6.9 times the interorbital width. Eyes and orbits moderately small and not elevated above level of cranium, orbits not well-defined, their estimated horizontal diameter 1.1 times interorbital width and 16% of preorbital snout length; interorbital width narrow, 3% of TL and 14.5% of preorbital snout length; spiracles small and circular, close posterior to eyes, their length 32% of orbit horizontal diameter; interspiracular space 2.0 times the interorbital width; 7 pseudobranchial folds in each spiracle. Tail cord-like, hardly tapering towards its tip, length from mid-vent to tip shorter (0.9 times) than body length from tip of rostral lobe to mid-vent; tail length 46% of TL; dorsal fins and lateral tail folds absent; tail width to height at level of pelvic tips 1.3; tail trapezoid in cross-section; epichordal caudal lobe probably confluent around tail tip, with shorter hypochordal lobe.
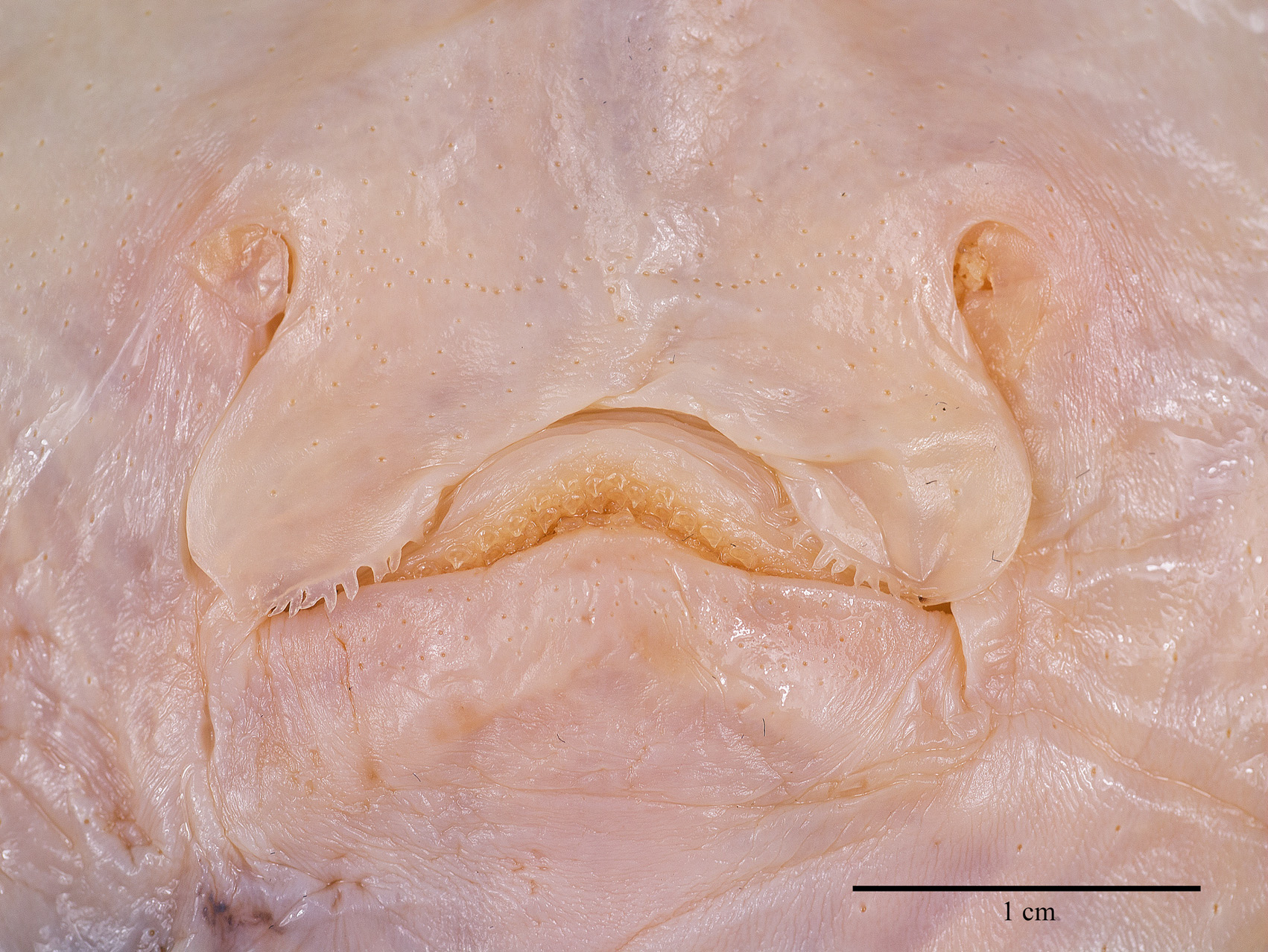
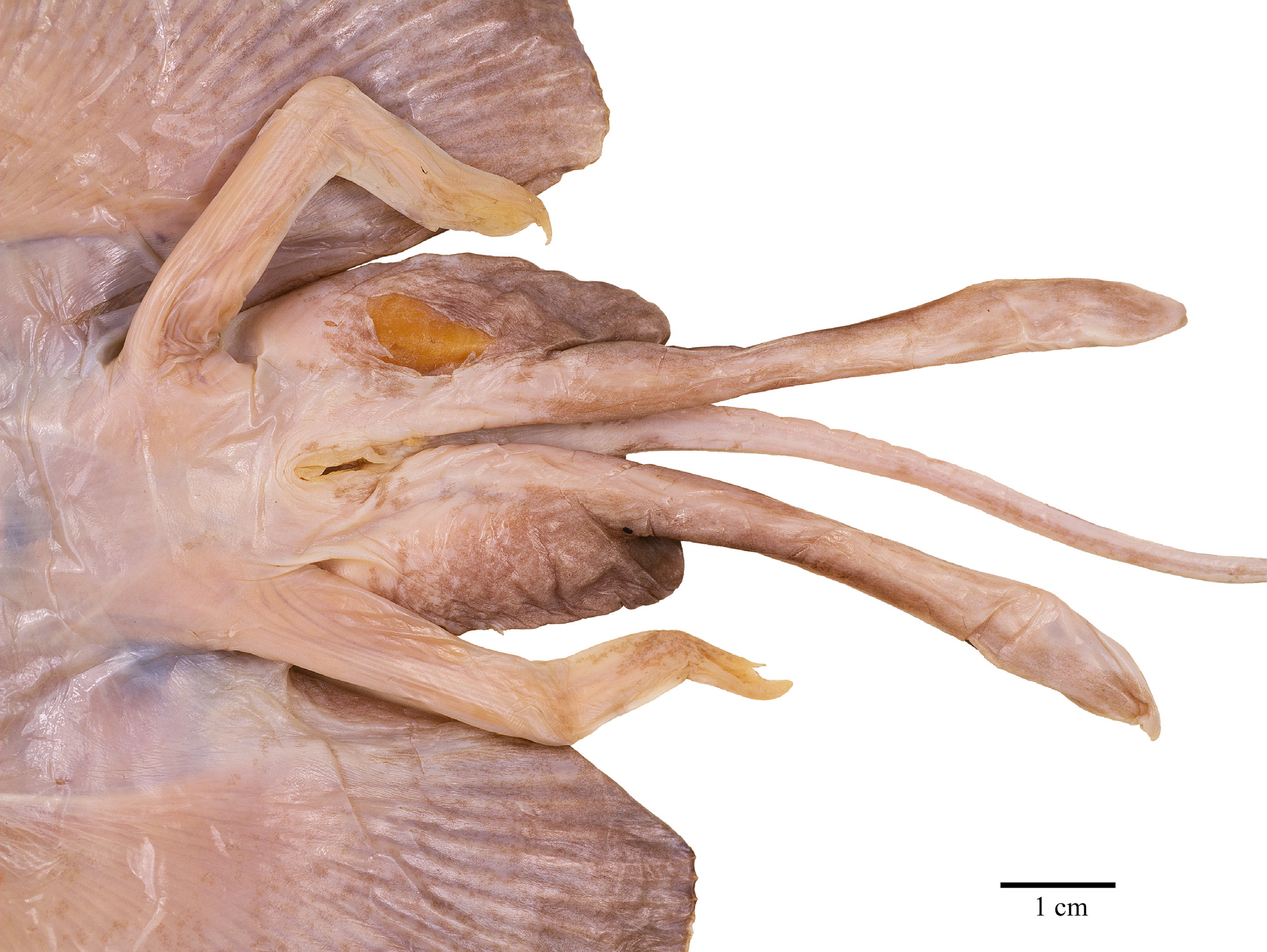
Ventral head length 34% of TL, 54.8% of disc length and 6.3 times the internarial width. Preoral snout length 23% of TL, 2.8 times in disc length and 4 times the mouth width. Mouth cavity appears to be set with indistinct papillae; mouth relatively small, jaws moderately angled; mouth width 16% of ventral head length and equal to internarial space; 24 tooth rows in upper and 22 in lower jaws, set in quincunx pattern; individual tooth with low, subcircular to rhombic base, tooth crown flat and with moderately long, stout, conical cusp. Gill openings small, length of first gill slit 0.8% of TL; distance between fifth gill slits 49% of distance between first gill slits, the latter 2.2 times the internarial space. Anterior nasal flaps low and fleshy, edges weakly serrated and undulated, terminal part set with short fringes; nasal curtain long and with papillose underside, oriented in longitudinal direction and laterally strongly protruding, outer edges broadly rounded, short and nearly transverse posterior margins set with short, stout, fleshy fringes; isthmus an evenly deep arc. Pelvic fin anterior lobes slender, leg-like, separated from and 0.9 times as long as posterior lobes; posterior lobes fin-like, their inner margins fused to base of tail along most of their length, with a short, pointed free tip. Claspers almost fully developed, very long, and moderately slender, with a slight constriction in the central part of the solid stem; terminal part relatively long and evenly tapering to tip; claspers exceeding tip of posterior pelvic lobes to almost half of tail length, their postanus length 50% of tail length.
mm % TL
total length (TL) 331.0 100.0
disc, width 212.0 64.0, length 208.0 62.8, length, point-to-point (ptp) 210.5 63.6, width at estimated ant. orbits 125.0 37.8
snout length, pre-eye 74.0 22.4, preorbital (estimated) 72.5 21.9, preorbital (est., ptp) 74.0 22.4, preoral 74.5 22.5, prenasal 68.3 20.6
orbit, horizontal diameter (est.) 11.5 3.5
eyeball, horiz. diameter 8.0 2.4
interorbital width 10.5 3.2
spiracle length, slit (opening proper) 3.7 1.1
interspiracular width 21.4 6.5
eye + spiracle length 14.8 4.5
orbit (est.) + spiracle length 16.7 5.0
Caudal fin (C), upper base length 26.0 7.9
Caudal fin (C), lower base length 19.0 5.7
tail, height at pelvic-tips 2.4 0.7, width at pelvic-tips 3.2 1.0
, height at upper C-origin 1.5 0.5
, width at upper C-origin 1.3 0.4
head length, ventrally 114.0 34.4, dorsally 90.8 27.4
mouth width 18.5 5.6
internarial width 18.2 5.5
nasal curtain, length 11.0 3.3, width each lobe 7.4 2.2
, space between lobes 9.0 2.7
gill slit length, 1st 2.8 0.8, 3rd 2.8 0.8
, 5th 2.0 0.6
interspace first gill slits 40.5 12.2 fifth gill slits 20.0 6.0
pelvic-fin length, ant. lobe 44.0 13.3, post. lobe 49.3 14.9
pelvic ant. lobe base width 10.0 3.0
......continued on the next page Sensory pore pattern. Few low, fleshy, tubular papillae dorsally on disc, concentrated particularly around the alar thorn fields; a row of widely spaced papillae proceeds rather diagonally from about level shoulder girdle to about median part of posterior pelvic lobes, tail with a lateral row of small tubular papillae to about level caudalfold origin, row missing in parts of anterior half of tail where it was probably abraded (remaining papillae in anterior half of tail bright, papillae in posterior half darker); a row of five pores close laterally alongside the base of the rostrum, an irregular row of densely set pores in anterior half of median trunk near the vertebral column, dorsal tail pores slit-shaped, but only partially detectable due to abrasion of the skin. Fields of densely arranged pores prenasally in basal half of rostrum, narrowly triangular fields of pores from nasal region outwards until half the distance to disc margins, a single irregular, transverse row of densely set pores internasally between anterior nasal flaps, and about two irregular, angled transverse rows in the skin covering the lower jaw.
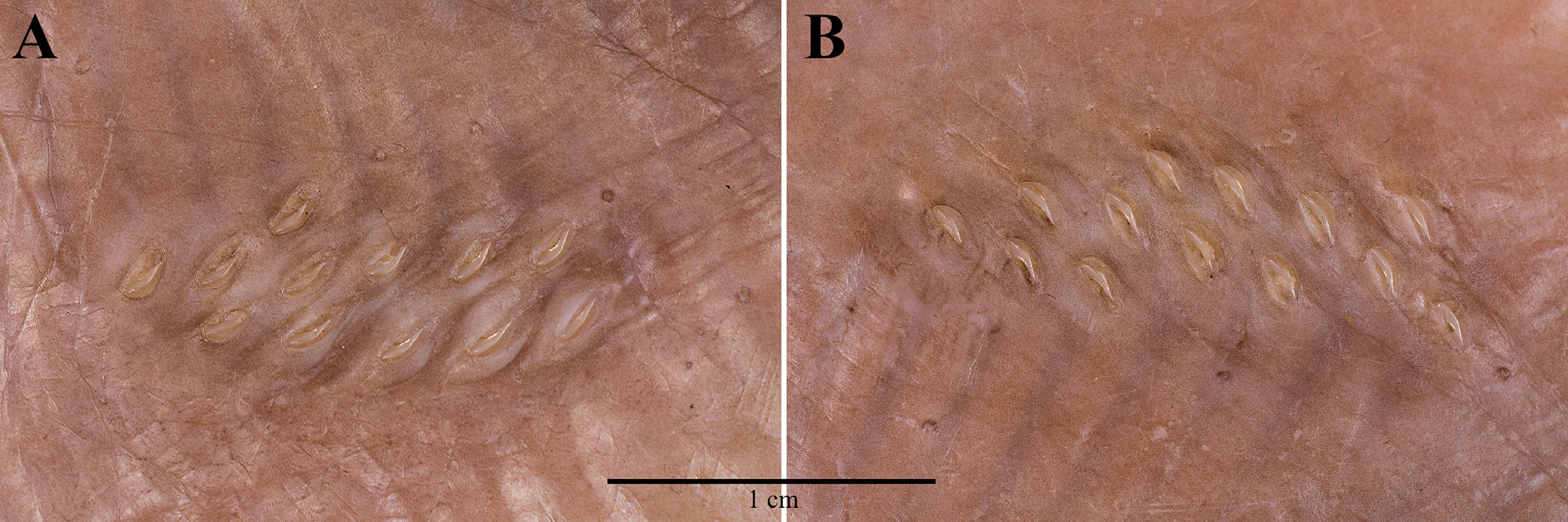
Squamation. Upper and lower surfaces of disc and tail entirely naked except for the alar thorns; alar thorns sharp, claw-like, not erectile, embedded in individual, transverse dermal pockets, and arranged in an elongated field of approximately two irregular longitudinal and six to seven transverse rows across wing tip.
Colouration (in preservative). Upper disc uniformly medium greyish-brown, brighter anterior to eyes in semitransparent side areas of brown rostrum; eyes dusky; anterior pelvic lobes distinctly lighter, beige; claspers dark as disc, except for laterally whitish terminal region. Ventral disc generally lighter than dorsal surface, beige with darker outer and posterior areas, slightly brighter in anterior half, and skin of belly cavity nearly transparent; nasal flaps and curtain creamy-white; mouth cavity whitish, teeth yellowish; pelvic fin posterior lobes medium greyishbrown like broadly posterior disc margins, anterior lobes distinctly brighter, beige. Claspers light greyish-brown, with medium greyish-brown origin and outer edges. Alar thorns distinctly marked off creamy-white in light grey pockets. Pores not distinctly marked, but with indistinct dark borders; tubular papillae marked blackish. Tail creamy-white ventrally, dorsally medium greyish-brown along most of its length, but terminal about 40 mm creamy-white with few brown blotches, caudal fold nearly transparent.
Claspers ( Fig. 12 View FIGURE 12 ). Almost mature, with all components formed, but skeleton still weakly calcified and somewhat flexible. Claspers long and slender, with stem of equal width but with slightly narrower width at about half clasper length; terminal region about one third of clasper length and only little widened, narrowing to pointed tip. Components on outer terminal surfaces absent. Inner dorsal terminal lobe without defined components, except for a slit (sl) across distal third transversally over a shallow cleft (cf) between axial and distal part of dorsal terminal 3 cartilages; axial cartilage running medially through entire terminal region as somewhat elevated ridge. Inner ventral terminal lobe in median third along outer edge with shield (sh) covered by smooth integument without lamellae and without outer free cartilage edge; its proximal pointed tip also hidden under integument and not forming free external component; its inner, nearly vertical wall distally forming external component dike (dk). The apparent component pent (pe) originating somewhat distally of proximal shield tip as a bulbous, fleshy bulge and running narrowing alongside inner edge of shield to near clasper tip; its outer surface not structured, but inner surface of the rolled-up component lamellated. Component sentinel (st) a sharply pointed free cartilage medially in distal third of ventral lobe, with two components sentina, of which the inner one (se1) spanning as integument cavity from inner origin of sentinel transversally onto the axial cartilage ridge, and the outer one (se2) diagonally between outer sentinel edge and distal parts of pent and dike. Component spike (sk) as a pointed free cartilage tip underneath sentinel and sticking out of se2 but exceeding sentinel length.
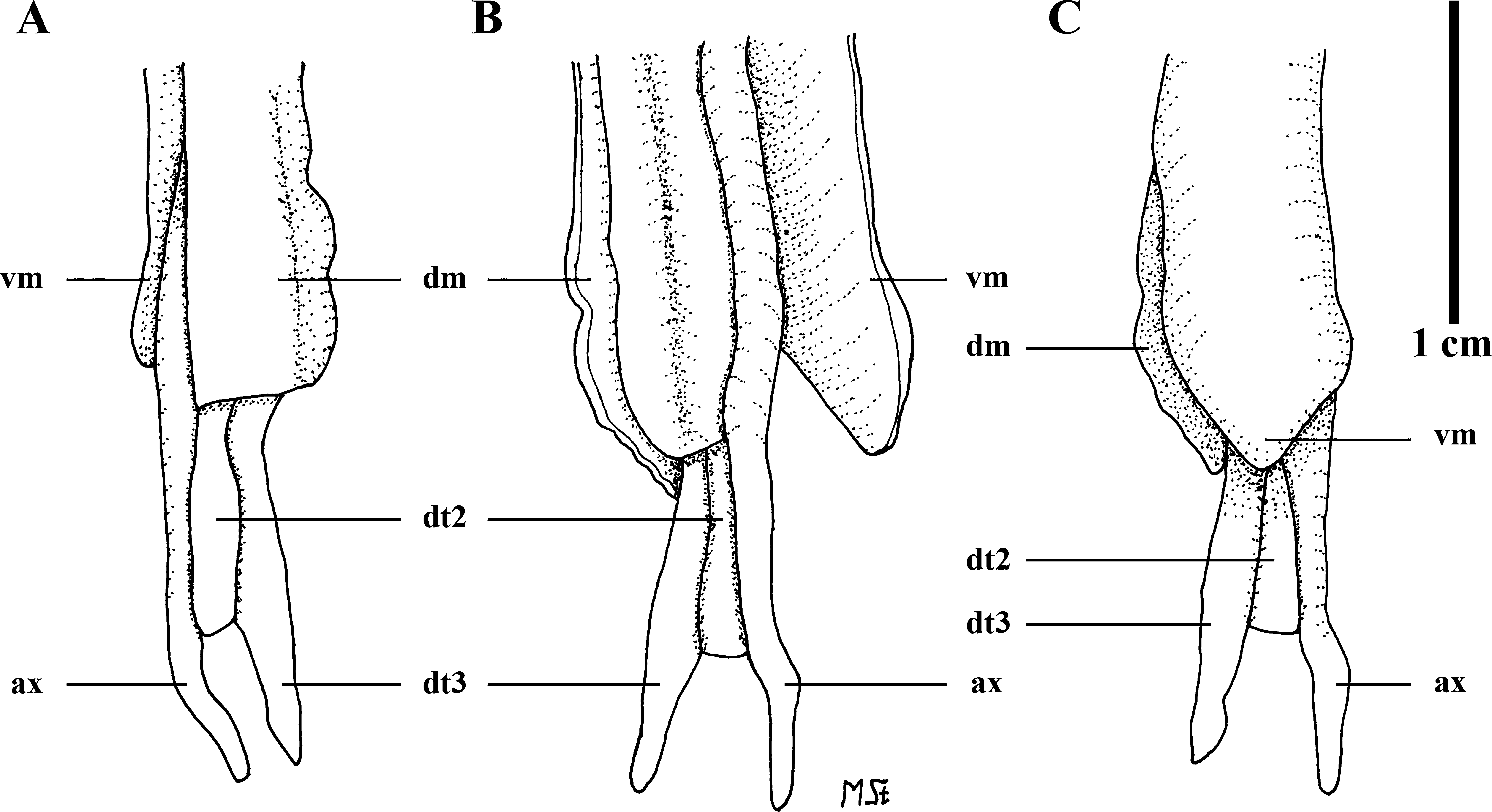
Clasper skeleton ( Figs. 13–16 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 ). Consisting of eight cartilages, namely axial (ax), dorsal (dm) and ventral (vm) marginals, two dorsal terminals (dt2+dt3), ventral terminal (vt), two accessory terminals (at1+at2). The right clasper skeleton was dissected on the specimen from somewhat proximal of terminal region, why further proximal parts of ax with beta-cartilage (β) and of both, dorsal and ventral marginals could be inspected only in radiographs. The β-cartilage on the head of ax is an exceptionally long, narrowly triangular element exceeding proximally the length of the pelvic fin's b3-element, which has no radial attachments, and reaching to about half length of the very short b2-element ( Fig. 13 View FIGURE 13 ). Proximal end of vm shorter than that of dm, and that of dm with truncate, that of vm with bluntly pointed edge. The ax is rather slender and running to the very end of the clasper skeleton, with weak calcification and somewhat curved, bluntly rounded distal tip. In dorsal view, the distal end of dm is truncate, with irregularly undulated outer edge in distal fourth of dm marked off as a thickened and somewhat separated margin ( Fig. 14 View FIGURE 14 ); this gives in x-rays the impression of a seemingly separate dorsal terminal 1 cartilage, and such a true separate dt1 has been found in Indobatis ori ( Weigmann et al. 2014) . However, a dt1 is absent in S. kotlyari clasper skeleton. In ventral view, the distal end of the vm is bluntly triangular, or spear-head shaped, with a weak constriction of both edges proximal of vm-tip ( Fig. 14 View FIGURE 14 ). Dorsal terminal 2 and 3 cartilages weakly calcified and not showing as external components on inner dorsal clasper lobe; both are elongated but dt2 is shorter with only two thirds length of dt3; dt2 with truncate distal end and fused with inner edge of dm and along its entire length with ax and dt3. The dt3 is more slender than dt2 and fused with distal edge of dm directly aside head of dt2; its distal end somewhat marked off by weak constrictions of both edges, with the tip pointed. Ventral terminal cartilage (vt) weakly calcified and elongated, twisted at half length and carrying proximally a solid, strongly inward curved process originating from the dorsal inner edge ( Fig. 15 View FIGURE 15 ); the outer proximal edge is distinctly serrated in both, ventral and dorsal views but not (possibly not yet?) appearing externally at outer edge of shield. The distal half of the ventral terminal is twisted to form an almost vertical wall as the external component dike. Like dt2 and dt3 for the dorsal clasper glans lobe, provides the vt the main cartilaginous support for the ventral lobe. Both elongated, slender accessory terminal cartilages are solid and well calcified; they are separately attached side by side to the very inner distal edge of the vm; the almost straight at2 is a little shorter than the in proximal half strongly undulated, or twisted at1 ( Figs. 13 View FIGURE 13 , 16 View FIGURE 16 ).
Snout skeleton and cranium ( Fig. 17 View FIGURE 17 ). Anteriormost pectoral skeleton elements falling short of rostral tip. Rostral cartilage extending to snout tip, very slender and appearing flexible though well calcified; its length more than twice length of neurocranium. Single cranial fontanelle very elongated, with broadly trapezoid posterior part with weakly concave transverse edge, gradually narrowing to level of anterior nasal capsules, but from there strongly narrowing to bluntly rounded tip; length of cranial fontanelle 39% of total cranial length and reaching forward into one fourth rostrum length. Maximum cranial width across nasal capsules 40% of total cranial length; maximum occipital width of neurocranium 75% of maximum cranial width; minimum dorsal interorbital width 30% of maximum cranial width.
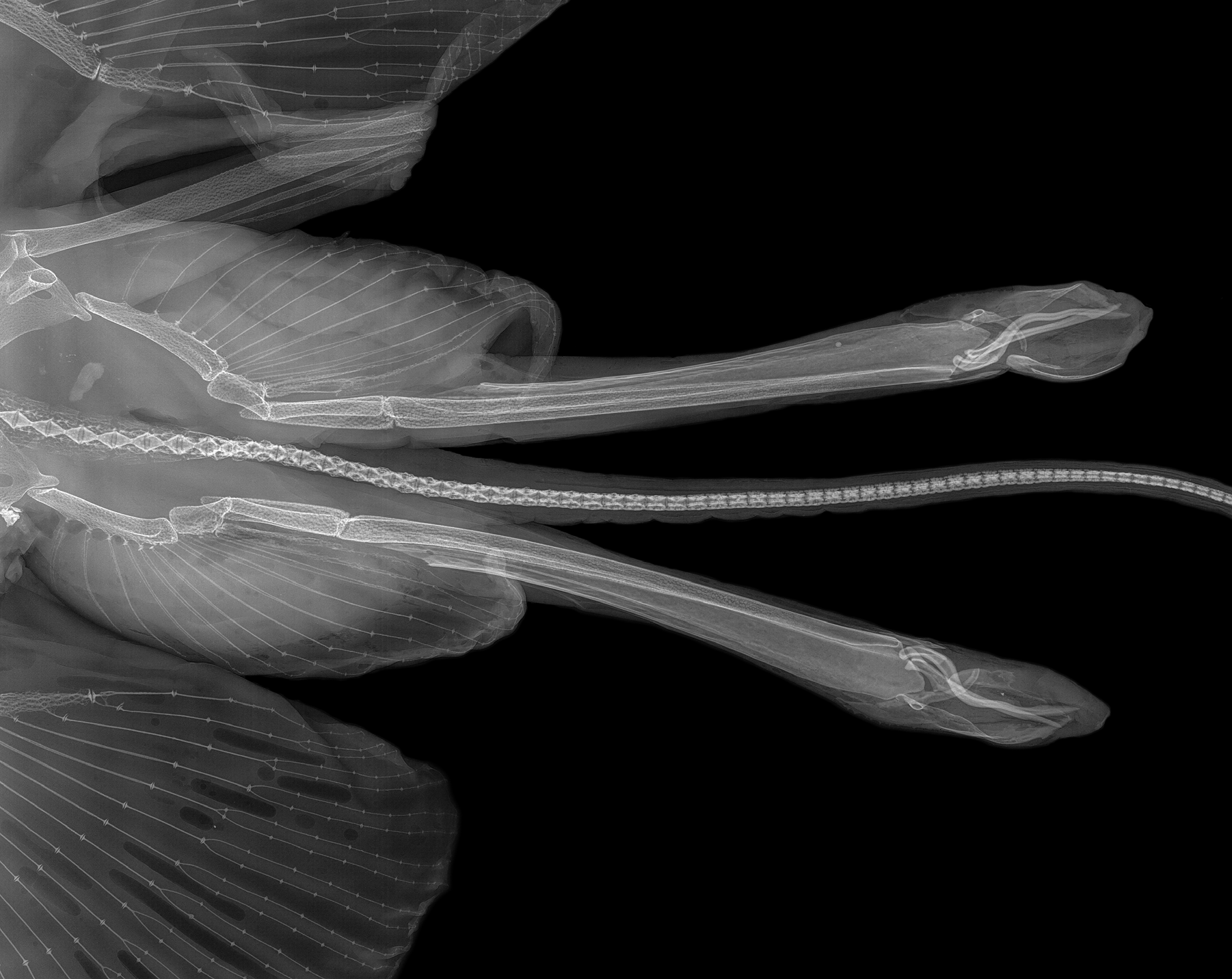
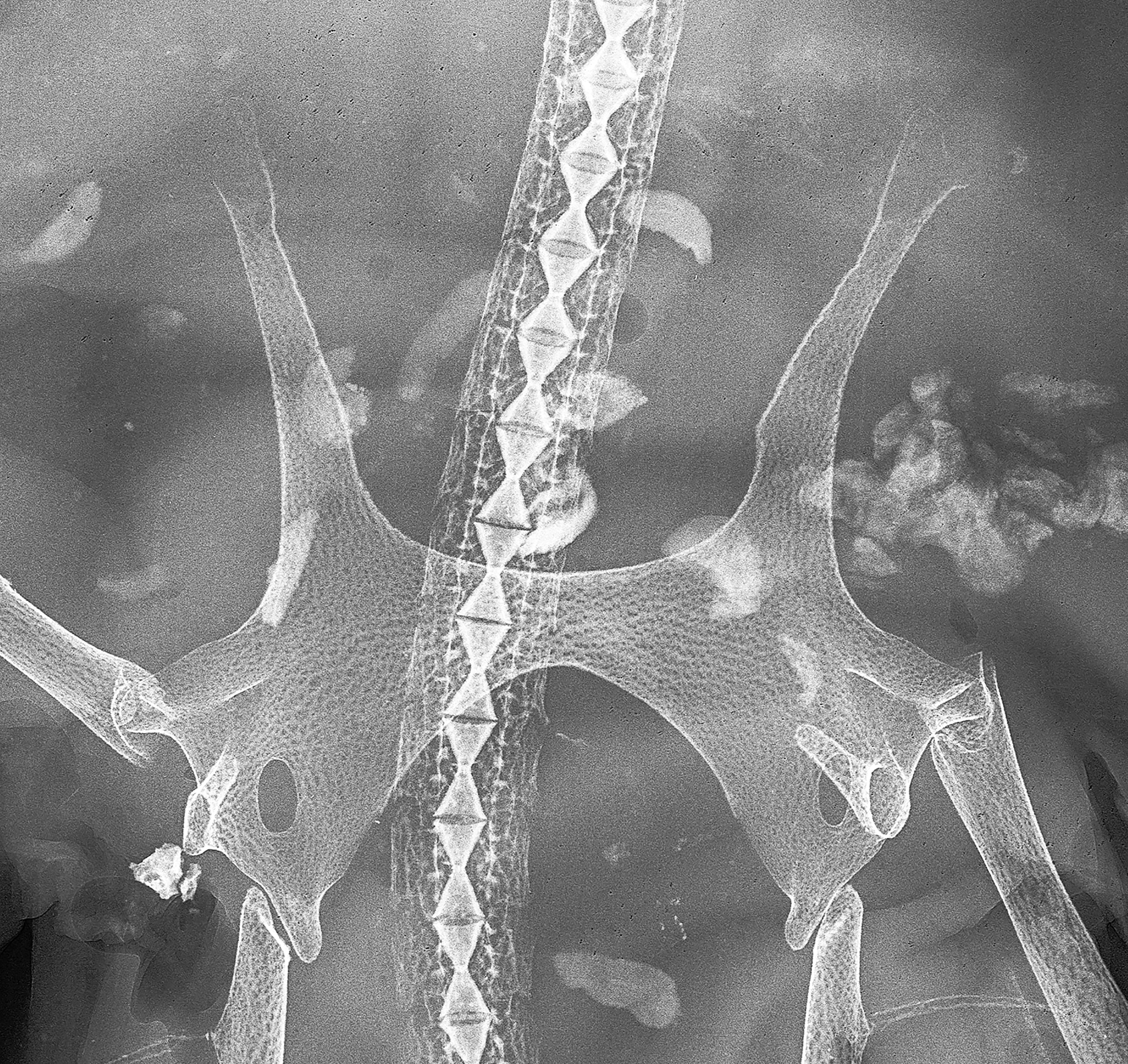
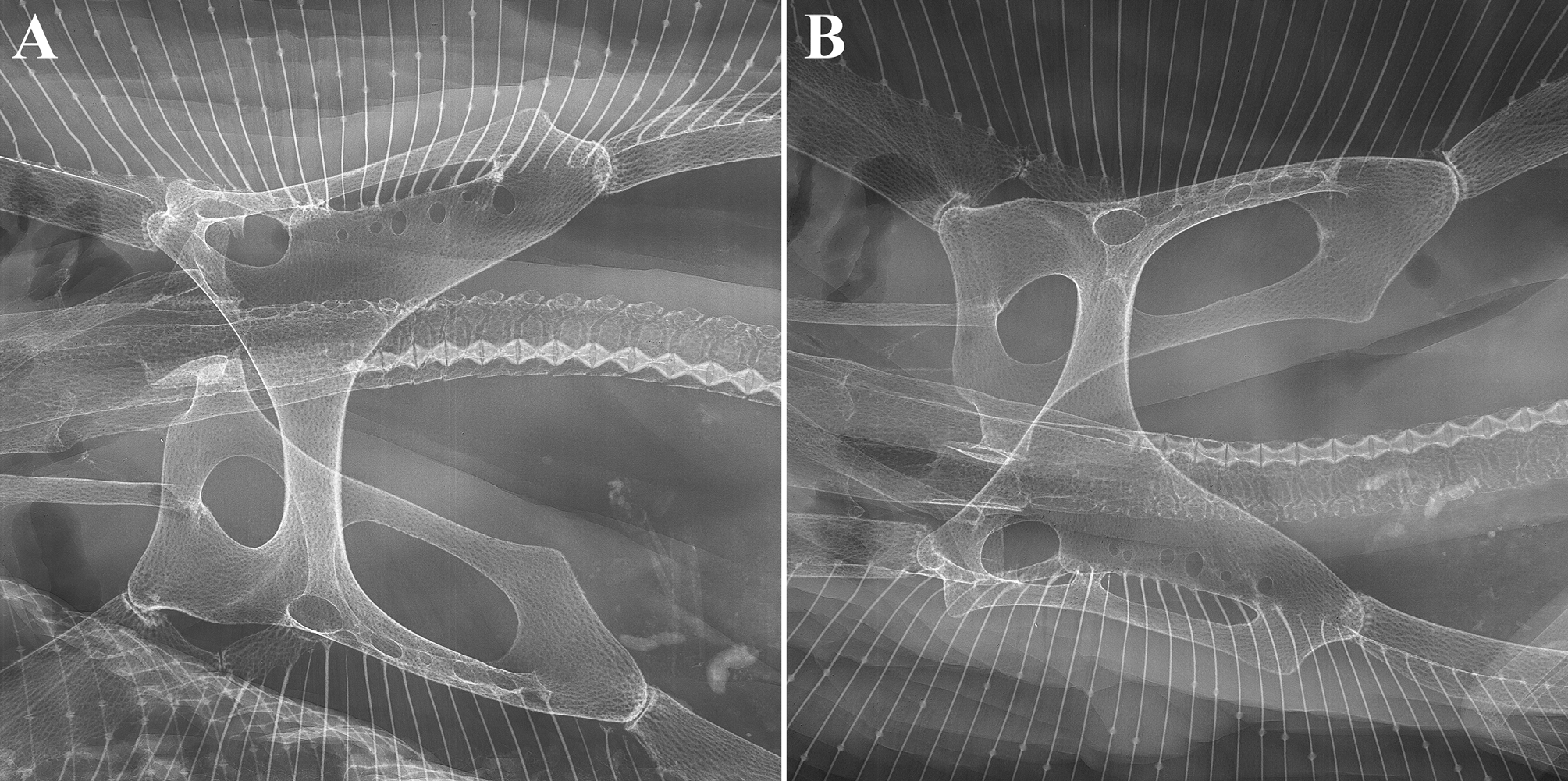
Pelvic girdle ( Fig. 18 View FIGURE 18 ). Rather narrow typical anacanthobatid element with massive iliac regions, which show each only one large foramen posteriorly; anterior edge of ischiopubic bar moderately concave, posterior contour a very deep, bluntly ovoid arc; prepelvic processes solid, very long and curved outward, their length from axis of maximum pelvic girdle width 75% of pelvic girdle width, about 54% measured from level of anterior contour pelvic girdle medially. Maximum width pelvic girdle 50% of maximum width shoulder girdle.
Scapulocoracoid ( Fig. 19 View FIGURE 19 ). Overall shape elongated rectangular and strongly asymmetrical regarding position of the mesocondyle; post-mesocondyle length nearly 2 times the pre-mesocondyle length. Moderately large anterior fenestra subcircular and not divided by anterior bridge; its position almost completely above longitudinal axis of scapulocoracoid. Single very large postdorsal fenestra of horizontally ovoid shape and a little more than 2 times the length of anterior fenestra. Six postventral foramina much smaller and in horizontal line below radial bearing post-mesocondyle ridge; first foramen the smallest and placed under the posterior edge of mesocondyle. Dorsal edge of the element weakly concave and gently sloping to well marked rear corner; posterior margin sloping concavely to metacondyle. Scapular process short and subquadrangular, with slightly oblique edge.
Shoulder girdle ( Figs. 17 View FIGURE 17 , 19–20 View FIGURE 19 View FIGURE 20 ). Radial bearing first proterygial basal element of pectoral fin skeleton inserting at mesocondyle much shorter than further anterior basal elements, about 4 times in length of the next anterior element, which is a divided cartilage as typical for male anacanthobatids (see Weigmann et al. 2014). First metapterygial basal element extremely long and bearing by far the majority of posterior pectoral radials; the very much shorter second and final metapterygium bearing only two radials laterally and two splitting from its posterior tip.
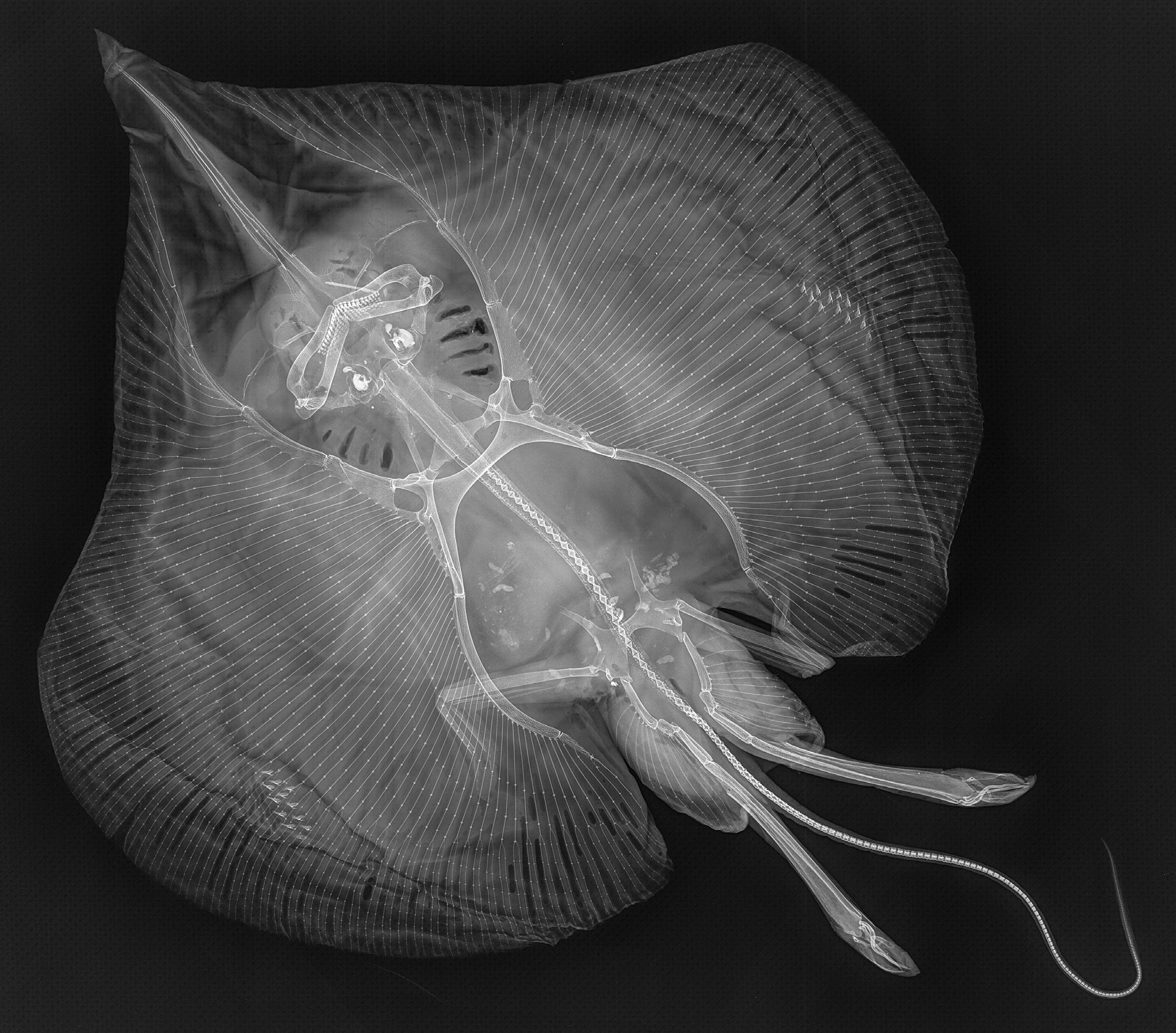
Skeletal meristics (from radiographs, Table 1, Fig. 20 View FIGURE 20 ). Free trunk vertebrae (Vtr) 27, plus only one vertebral centrum within synarcual; tail vertebrae ~173, total vertebrae ~200; pectoral radials left/right 75/75, pelvic radials left/right 3+12/3+12.
Size. A small species of Sinobatis reaching at least 331 mm TL. The male holotype is almost adult as its claspers have apparently reached their maximum size and all clasper components and alar thorns are developed. However, the cartilages are still not fully calcified and somewhat flexible. Therefore, it is assumed that males mature at around 33 cm TL. Based on a comparison of the relation between maturity size and maximum size in other legskate species, a maximum size of about 42 cm TL is assumed for the new species. This putative maximum size is based on a mean exceedance of maximum size vs. maturity size of 26% in legskates, calculated from data of maturity and maximum sizes of Anacanthobatis marmorata (maturity size 23 cm, maximum size 29 cm), A. americana (31 cm, 38 cm), Sinobatis borneensis (25 cm, 35 cm), S. bulbicauda (43 cm, 56 cm) and S. melanosoma (53 cm, 60 cm) from Séret et al. (in press). Adult females may grow to even somewhat larger size, as is well documented for many rajiform species.
Legend to symbols: + = present/yes; - = absent/no; +- = present or absent/yes or no; ++ = two times present; ++- = two times present or absent; +/++ = one or two times present
Distribution ( Fig. 1 View FIGURE 1 ). The new species is known only from the remote Broken Ridge in the southeastern Indian Ocean where the holotype was caught in 1400 m depth.
Etymology. Named in honour of Dr. Alexander Kotlyar, IOAN Moscow, who collected and preserved all fish samples during the cruise 37 years ago in 1979, when he was working at VNIRO Moscow. He forwarded all samples to ZMMU for deposition and detailed study. Without his kind assistance, it would hardly have been possible to reconstruct and verify from his original logbook the station data of the holotype.
Remarks. In radiographs ( Fig. 18 View FIGURE 18 ), the prepelvic processes of the new species show the seeming terminal bifurcation that was reported by Weigmann et al. (2014) and Weigmann & Stehmann (2016) for several other anacanthobatids, apparently caused by limited depictability due to weak calcification in the central parts of the tips.
The type locality at Broken Ridge is very remote and might be indicative of local endemism. A similar case of a new Sinobatis sp. with remote and restricted distribution has been found with a species from the Saya de Malha Bank in the central western Indian Ocean (Weigmann & Stehmann 2016). So far, the western Pacific, western Atlantic and western Indian Oceans have been identified as hotspots for anacanthobatids with the highest species numbers (Weigmann 2016). The new Sinobatis species described herein, however, already is the third species of the family known from the eastern Indian Ocean, indicating that the eastern Indian Ocean might represent another hotspot.
For a key to the species of Anacanthobatidae in the Indian Ocean, including Sinobatis kotlyari n. sp. (as Sinobatis sp. Broken Ridge), see Weigmann & Stehmann (2016).
Resurrection of the family Gurgesiellidae de Buen, 1959, including Cruriraja Bigelow & Schroeder, 1948 , Fenestraja McEachran & Compagno, 1982 and Gurgesiella de Buen, 1959 , is planned based on morphological and molecular data (Weigmann 2016; Weigmann et al., in press).









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.