
Anisuracampa ywangana Sendra & Komerički, 2021
|
publication ID |
https://doi.org/10.5852/ejt.2021.731.1199 |
|
publication LSID |
lsid:zoobank.org:pub:A734B727-9978-4E14-B431-85865CE91E40 |
|
DOI |
https://doi.org/10.5281/zenodo.4423298 |
|
persistent identifier |
https://treatment.plazi.org/id/511F81B5-6CB2-4A98-AB3E-11AF72205A26 |
|
taxon LSID |
lsid:zoobank.org:act:511F81B5-6CB2-4A98-AB3E-11AF72205A26 |
|
treatment provided by |
Plazi (2021-01-07 08:28:06, last updated 2024-11-26 09:02:22) |
|
scientific name |
Anisuracampa ywangana Sendra & Komerički |
| status |
sp. nov. |
Anisuracampa ywangana Sendra & Komerički sp. nov.
urn:lsid:zoobank.org:act:511F81B5-6CB2-4A98-AB3E-11AF72205A26
Figs 1, 3–19 View Figs 1‒4 View Figs View Figs 9‒11 View Figs 12‒15 View Figs 16‒19 ; Tables 1–2 View Table 1 View Table 2 ; Supp. file 2
Etymology
This species is named after the important karst region Ywangan in Myanmar, the location of Win Twin Cave ( Figs 1, 3–4 View Figs 1‒4 ).
Type material
Holotype
MYANMAR • ♀; Shan State, Ywangan, Win Twin Cave ; 21º11′51.47″ N, 96º32′40.34″ E; 16 Nov. 2018; Aung Lin leg.; labelled “holotype”; MZB ( MCNB) 2020-1152 .
GoogleMapsParatypes
MYANMAR • 1 ♀; same collection data as for holotype; labelled “paratype-H1”; MZB ( MCNB) 2020- 0620 GoogleMaps • 1 ♀, 1 ♂, 1 juvenile; same collection data as for holotype; 8 May 2018; labelled “paratype-H2”, “paratype-M”, “paratype-J”; MZB ( MCNB) 2020-1153 , MZB ( MCNB) 2020-1155 , MZB ( MCNB) 2020-1154 GoogleMaps .
Other material examined
MYANMAR • 2 specs; same collection data as for the holotype; 8 May 2018; mounted on two separate aluminium stages and coated with palladium-gold; Coll. AS. GoogleMaps
Description
BODY. Length is 4.3–6.0 mm (n = 4) in adults and 2.6 mm in one juvenile ( Table 1 View Table 1 ). The epicuticle is smooth under optical microscope but reticulated at higher magnification, showing irregular polygonal structures of variable sizes ( Fig. 11 View Figs 9‒11 ). Body with long smooth clothing setae.
HEAD. Two complete and intact antennae in holotype and paratype-M with 35 antennomeres each, whereas paratype-J (juvenile) has 38 antennomeres. The antennae are similar to length of body ( Table 1 View Table 1 ), with medial antennomeres 2× as long as wide but apical antennomeres 1½ × as long as wide. Cupuliform organ with up to 17 spheroidal olfactory chemoreceptors arranged in two uneven concentric circles; each chemoreceptor forms a complex structure of multi–perforated folds with one distinguishing crown of fringes a surrounding central column. Each of these structures is inside a polygonal cell ( Figs 5–6 View Figs ). Distal and central antennomeres with three whorls of distal barbed macrosetae and 5–6 scattered whorls of smooth setae in addition to single distal whorl of up to 14 gouge sensilla of 24–28 µm length ( Figs 7–8 View Figs ) that are more abundant on dorsal side of antennomere. Proximal antennomeres with typical trichobothria, plus small bacilliform sensillum on third antennomere in ventral position. Round protrusion of frontal process covered with one anterior macroseta and two or three posterior macrosetae ( Fig. 10 View Figs 9‒11 ). Three macrosetae along each side of insertion line of antennomere and x setae with thin distal barbs; length ratios of a / i / p / x, 45/57/56/65, respectively, in paratype-H2. Labial palps large and suboval, each with bacilliform latero-external sensillum, two guard setae, up to 18 normal setae and up to 140 neuroglandular setae.
THORAX. Thoracic macrosetae distribution ( Figs 9, 11 View Figs 9‒11 ) have pronotum with 1+1 ma, 1+1 la, 2+2 lp 2,3 macrosetae; mesonotum with 1+1 ma, 2+2 la and 4+4 lp 1–4 macrosetae; and metanotum with 1+1 ma, 2+2 la and 3+3 lp 1–3 macrosetae. All macrosetae long and with thin barbs along basal half to twothirds of each seta; marginal setae barbed and longer than clothing setae. Legs elongated, metathoracic legs reach end of abdomen in adults or overpass it in juvenile. Tibia always longer than femur or tarsus ( Table 1 View Table 1 ). Each femur III with three or, less frequently, two long dorsal macrosetae with distal barbs (0.25 mm in 0.62 mm femur of paratype-H1) (femora I–II with one to three long dorsal macrosetae). Calcars covered with thin abundant barbs all over. Tibiae I–III usually with two short, completely barbed ventral macrosetae, occasionally just one. Each tibia with two rows of ventral barbed setae almost from base and scattered throughout with thin, long setiform sensilla. Three well-barbed dorsal distal tarsal setae longer than rest of tarsal setae. Subequal claws with lateral expansion in crest; basal and ventral portion of claws covered with short to long spiniform formations. Laminar pretarsus of lateral processes sharply curved with what apparently looks like thin fringes under optical microscope, but under SEM seen as narrow laminar expansions ( Figs 12–15 View Figs 12‒15 ).
ABDOMEN. Distribution of macrosetae on tergites with 1+1 post 1 on I–III; 1+1 post 1 to 3+3 post 1–3 on IV, 4+4 post 1–4 and 1+1 la on V, 5+5 post 1–5 or 6+6 post 1–6 and 1+1 la on VI–VII; 7+7 post on VIII; and 8+8 or 9+9 post macrosetae on IX abdominal segment. All tergal abdominal macrosetae long and barbed along distal half to four-fifths. Urosternite I has 7+7 macrosetae ( Figs 16–17 View Figs 16‒19 ); while urosternites II to VII with 5+5 macrosetae; and urosternite VIII with 1+1 macrosetae; urosternal macrosetae of medium size and barbed along distal half. Stylus with short apical seta with small barbs, and each subapical seta completely covered with barbs and with short, barbed ventromedial seta ( Figs 18–19 View Figs 16‒19 ). One intact cercus 2.4 × body length and with 14 primary articles ( Table 2 View Table 2 ). Article length increases from proximal to distal articles; covered with long thin macrosetae, with distal barbs, and less abundant long, thin setae. Each primary article with whorl of shorter thin plumose setae at distal position.
SECONDARY SEX CHARACTERS. Female urosternite I with enlarged subcylindrical appendages, each bearing up to 60 ventral glandular a 2 setae proximally and apical field of up to 28 glandular a 1 setae ( Figs 16–17 View Figs 16‒19 ). Male with similar appendages with two fields of glandular setae (19 a 2 and 27 a 1) in one appendage of male paratype. Spermatozoid fascicles small, 0.04 mm in diameter, and formed by undistinguishable filament of spermatozoids in spiral.
Taxonomic affinities
Anisuracampa was proposed for a soil-dwelling species, Anisuracampa suoxiensis Xie & Yang, 1990 from Hunan Province in southeastern China. As a plusiocampine genus, Anisuracampa is characterized by the laminar pretarsus but with the lateral processes with long barbs and weakly developed lateral crests ( Figs 12–15 View Figs 12‒15 ) (see Xie & Yang 1991: fig. 34). In addition to this trait, Anisuracampa has two or three dorsal femoral macrosetae and 1+1 macrosetae on the eighth urosternite. All of these characters are present in A. ywangana sp. nov., a cave-adapted species from the Win Twin Cave in eastern Myanmar.
Therefore, these morphological similarities together with the congruent geographical distribution in Southeast Asia are considered sufficient reason to place this new species in Anisuracampa . However, A. ywangana sp. nov. differs from A. suoxiensis in several characters besides the traits related to adaptations to cave ecosystems. Anisuracampa ywangana sp. nov. has 4+4 lateral posterior macrosetae on the mesonotum ( Fig. 9 View Figs 9‒11 ) (2+ 2 in A. suoxiensis ), 3+3 lateral posterior and 2+2 lateral anterior macrosetae on the metanotum (2+2 lateral posterior and no lateral anterior macrosetae in A. suoxiensis ) and 5+5 posterior and 1+1 lateral anterior macrosetae on urotergites V–VII (4+4 posterior and no lateral anterior macrosetae in A. suoxiensis ). Regarding the new species’ cave-adapted features, it has a larger body and longer appendages at least double the size of those of A. suoxiensis , with 30–35 antennomeres in A. ywangana sp. nov. (24 in A. suoxiensis ). It is impossible to compare other useful taxonomical characteristics mentioned in the description of A. ywangana sp. nov. with A. suoxiensis as the former was described using SEM, whereas the latter was described under optical observations with a brief diagnosis and the type material of A. suoxiensis is not available for study.
Remarks
Anisuracampa ywangana sp. nov. was observed and collected walking on the Win Twin cave floor and boulders, approximately 300 m from the entrance, and on the wet flowstone in a vast chamber ( Figs 1–3 View Figs 1‒4 ; Supp. file 2). The Win Twin cave is located within a large karst area referred to as Ywangan karst, which is situated in the western part of the Shan plateau, approximately 10 km northeast of Ywangan Township and 15 km east of the Panlaung Pyadalin Wildlife Sanctuary. The cave has only been discovered recently and is currently under tourist development by the local community. It has not been fully surveyed and approximately only the first 400 m were explored during the collection of the type material. The cave itself is large, with numerous chambers, and its entrance was enlarged by mining ( Fig. 4 View Figs 1‒4 ); after approximately 300 m into the main passage, it opens into a vast chamber filled with speleothems ( Fig. 3 View Figs 1‒4 ), from which it again continues further by smaller, narrow passages opening into new chambers. At the bottom of the vast chamber, the oxygen level becomes too low for further exploration during the dry season (May), while by the end of the rainy season (November) it was possible to reach two further lower chambers before the oxygen level became too low. No active water flow was observed, but sparse remains of particular organic matter are present and the air temperature recorded in May 2018 was 20.0°C, while the sediment temperature was 19.6°C (unpublished data).
Xie R. & Yang Y. 1991. Description of two new genera and three new species of Campodeidae in China (Diplura). Contribution Shanghai Institute of Entomology 10: 95 ‾ 102.

Figs 1‒4. Cave-adapted diplurans from Win Tin Twin Cave, Ywangan karst, Shan State, Myanmar. 1. Anisuracampa ywangana Sendra & Komerički sp. nov., with Diplopoda at 300 m from the entrance. 2. Mueggejapyx brehieri Sendra & Komerički gen. et sp. nov. at the same site. 3. Speleothems in the main passage of the cave. 4. Entrance to the cave. (Photographs: Ana Komerički).
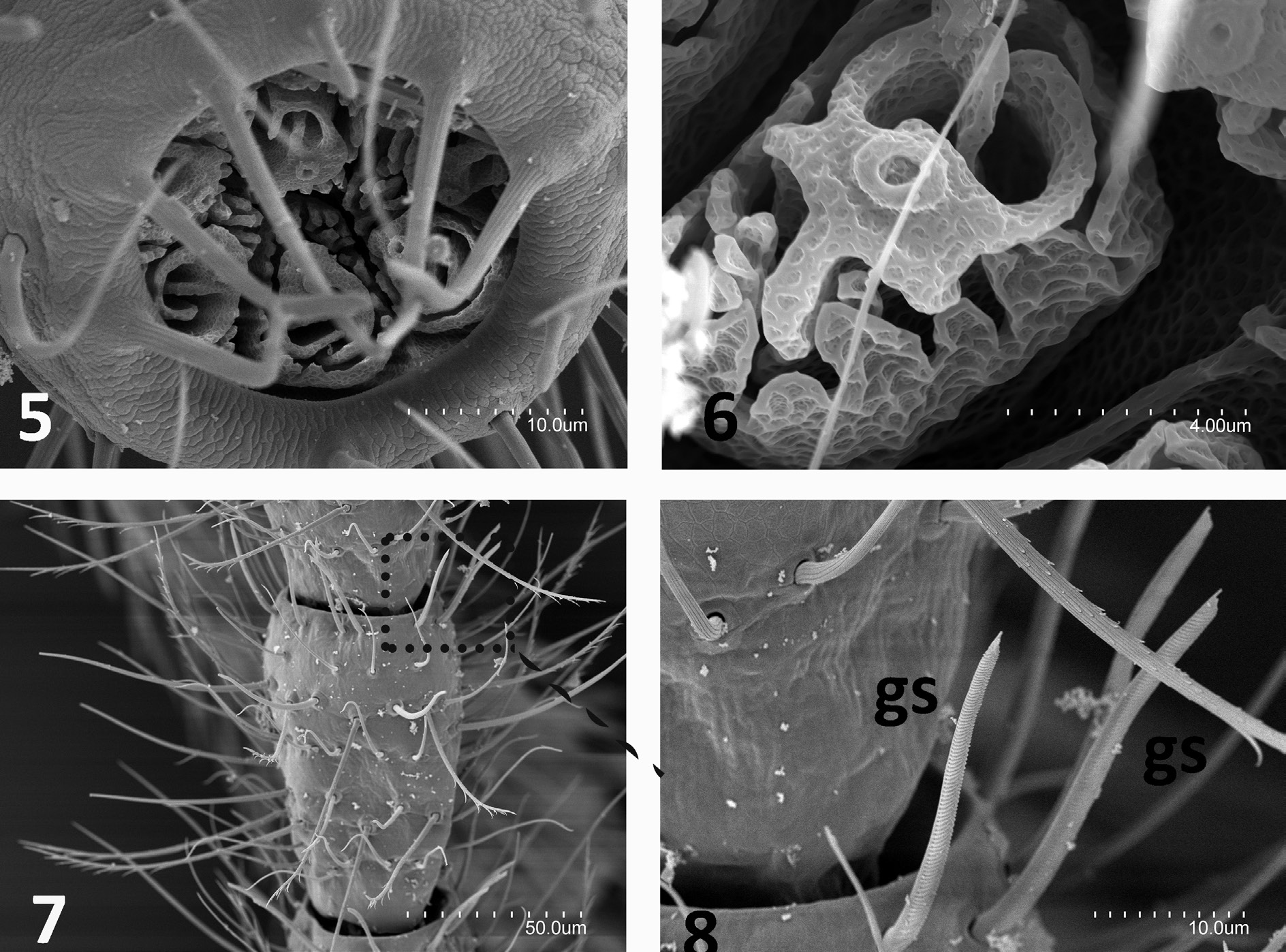
Figs 5‒8. Anisuracampa ywangana Sendra & Komerički sp. nov. 5. Apical view of last antennomere. 6. Olfactory chemoreceptor of the cupuliform organ. 7. Central antennemeres. 8. Lateral distal medial antenomere with gouge sensilla (gs).
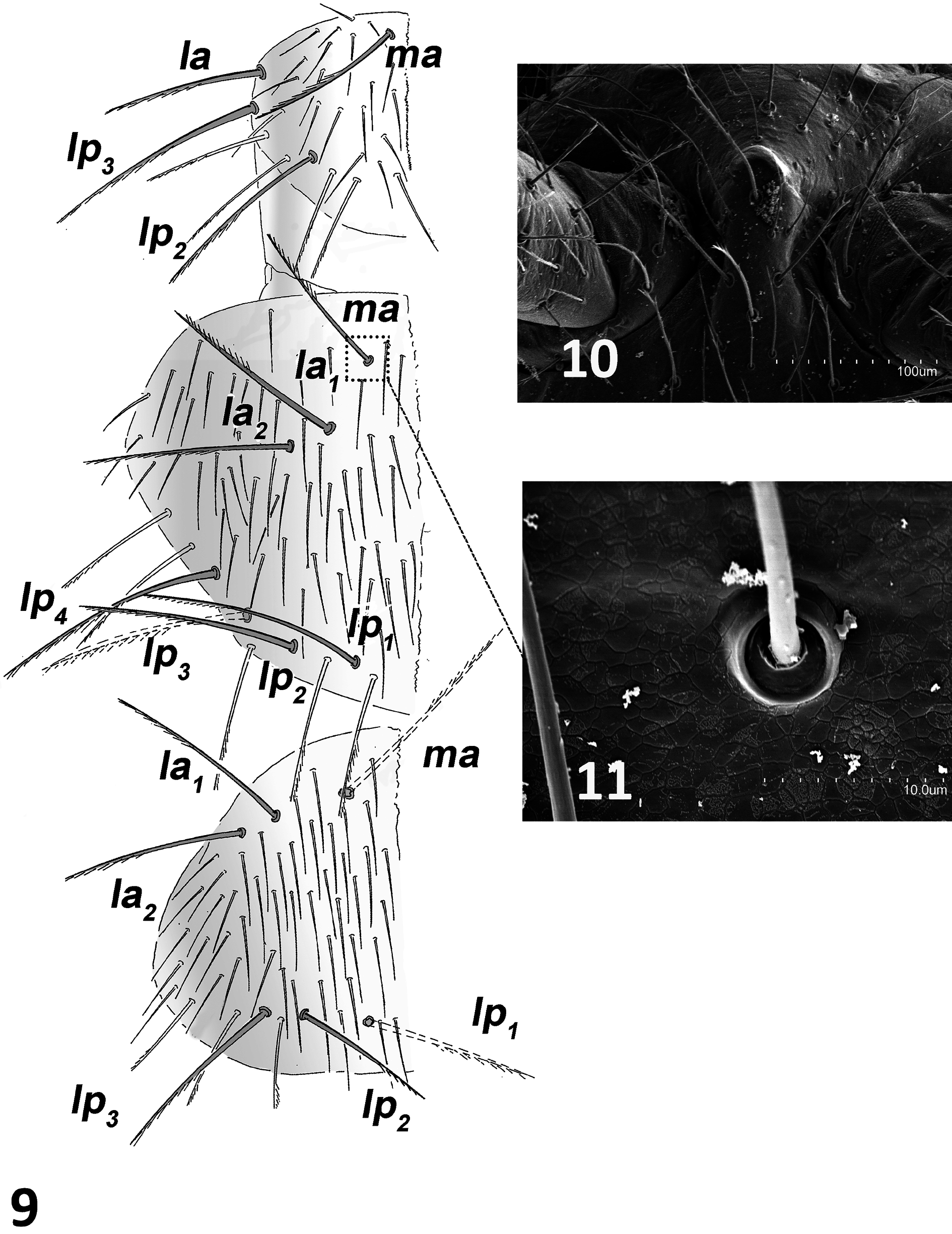
Figs 9‒11. Anisuracampa ywangana Sendra & Komerički sp. nov., holotype (MZB (MCNAB) 2020- 1152) 9. Pronotum, mesonotum, and metanotum, left side. 10. Frontal process. 11. Detail of the base of medial anterior mesonotal macroseta.
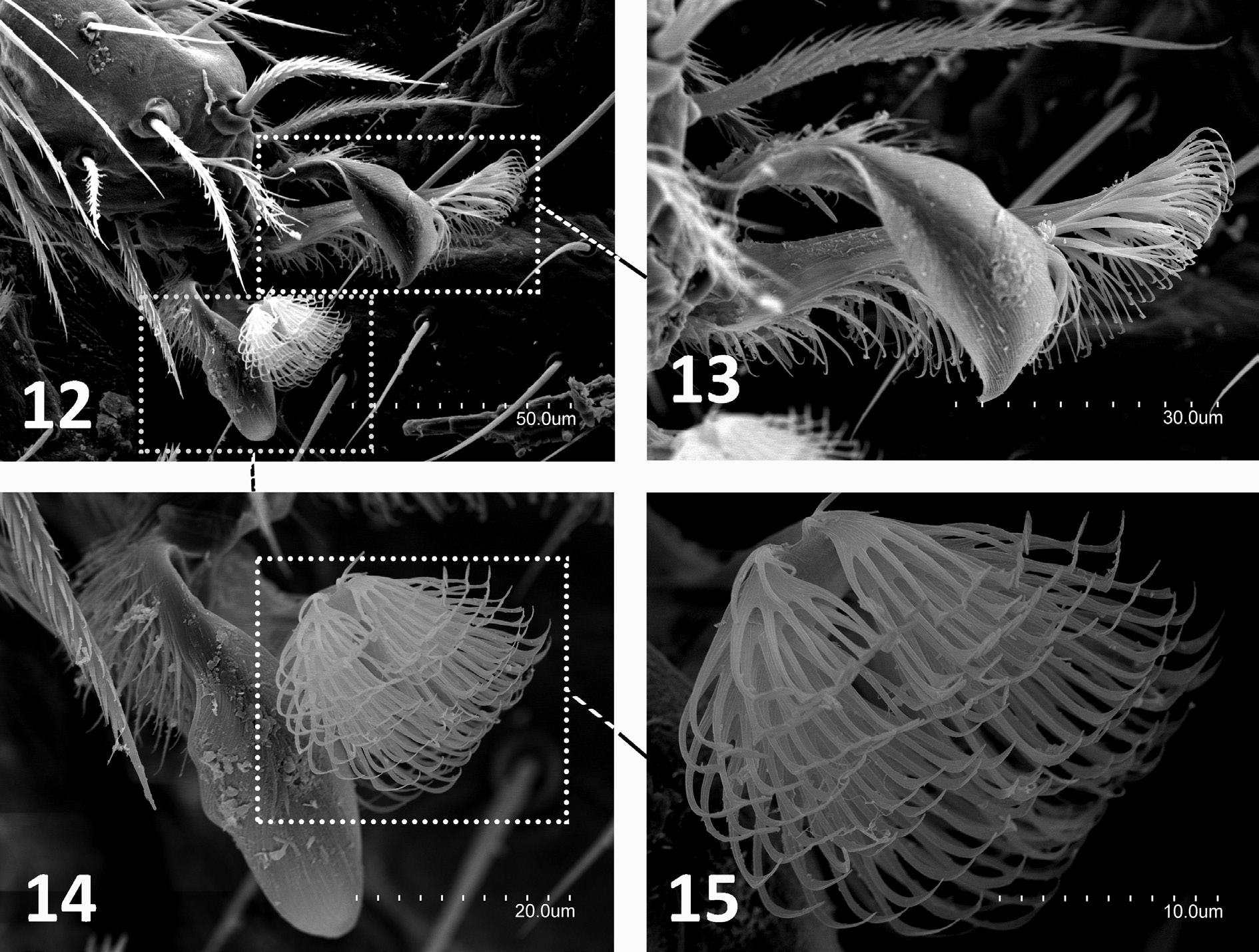
Figs 12‒15. Anisuracampa ywangana Sendra & Komerički sp. nov. 12. Pretarsus of metathoracic leg. 13. Posterior claw and its lateral processes of pretarsus. 14. Anterior claw and its lateral processes. 15. Detail of lateral processes.

Figs 16‒19. Anisuracampa ywangana Sendra & Komerički sp. nov. 16. First urosternite in a female. 17. Detail of the appendage of the first urosternite in a female with glandular a 1 and a 2 setae. 18. Stylus of the lateral right side of the fifth urosternite. 19. Detail of the stylus of the lateral right side of the fifth urosternite.
Table 2. Anisuracampa ywangana Sendra & Komerički sp. nov., paratype MZB (MCNB) 2020-1153, length of cercal articles and total length (units in mm), including number of articles of the cercus, basal article included, and division of basal article in secondary articles.
| Division- basal article | Basal article | 1 st | 2 nd | 3 rd | 4 th | 5 th | 6 th | 7 th | 8 th | 9 th | 10 th | 11 th | 12 th | 13 th | 14 th | Total cercus | Number of articles |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 1.1 | 0.39 | 0.45 | 0.50 | 0.55 | 0.65 | 0.75 | 0.83 | 0.92 | 0.94 | 1.20 | 1.44 | 1.65 | 1.67 | 1.64 | 14.68 | 15 |
| MZB |
Museum Zoologicum Bogoriense |
| MCNB |
Museu de Ciències Naturals de Barcelona |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Hexapoda |
|
Class |
|
|
Order |
|
|
SubOrder |
Rhabdura |
|
Family |
|
|
SubFamily |
Plusiocampinae |
|
Genus |
1 (by plazi, 2021-01-07 08:28:06)
2 (by ExternalLinkService, 2021-01-07 08:36:39)
3 (by valdenar, 2021-01-07 12:49:26)
4 (by valdenar, 2021-01-07 12:52:38)
5 (by ExternalLinkService, 2021-01-07 13:03:40)
6 (by ExternalLinkService, 2021-09-19 03:31:46)
7 (by ExternalLinkService, 2021-10-20 03:46:13)
8 (by plazi, 2023-11-01 15:37:24)
9 (by ExternalLinkService, 2023-11-02 09:57:01)