
Gobiodon fuscoruber, Herler, Juergen, Bogorodsky, Sergey V. & Suzuki, Toshiyuki, 2013
|
publication ID |
https://dx.doi.org/10.11646/zootaxa.3709.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:6F6CC130-6A51-4FC0-8E3C-A82B598FFB49 |
|
persistent identifier |
https://treatment.plazi.org/id/03E18789-1931-AD41-3AEB-FE31EABAFDA1 |
|
treatment provided by |
ImsDioSync |
|
scientific name |
Gobiodon fuscoruber |
| status |
sp. nov. |
Gobiodon fuscoruber , sp. nov.
Brown-red Coralgoby
Figs. 7 View FIGURE 7 , 8 View FIGURE 8 , 10 View FIGURE 10 & 11 View FIGURE 11 ; Tables 1, 8, 9 & 10
Gobiodon unicolor (non Castelnau, 1873) Munday et al. (1999): 56, Fig. 13; Hayashi & Shiratori (2003): 63, Fig. 116; Senou et al. (2004): 170; Harold et al. (2008): 132.
Gobiodon sp. 5 Akihito et al. (2002): 1190.
Gobiodon sp. D Senou et al. (2004): 175.
Gobiodon sp. 3 Herler and Hilgers (2005): 123, Fig. 15; Niedermüller et al. (2009): 1501, Fig. 2 View FIGURE 2 ; Herler et al. (2009): 733, Fig. 4 View FIGURE 4 .
Holotype. NMW 95079 View Materials (cited erroneously as CH 232 - 42 -003 (correct number: CH 232 - 43 -003) in Herler & Hilgers, 2005), male 36.7 mm SL, Gulf of Aqaba, Egypt, Dahab, “Islands” (28 ° 28 ’ 38.50 ” N, 34 ° 30 ’ 47.10 ” E), 1.5 m, coll. J. Herler, 27 March 2004.
Paratypes. Seven specimens. NMW 95080 View Materials , 31.3 mm SL, 3 m, 11 November 2005, other data same as holotype. MNHN 2006 – 1700, 32.1 mm SL, 2 m, 17 November 2005, other data same as holotype. BMNH 1951.1. 16.555, 29.2 mm SL and BMNH 1951.1. 16.556, 34.0 mm SL, Red Sea, Saudi Arabia, Sanafir Island, coll. N. Marshall (Manihine Expedition), 1951. OMNH 39984, female, 29.2 mm SL, Japan, Ryukyu Islands, Iriomote Island, coll. T. Suzuki, 0 7 August 1993. OMNH 39986, male, 23.6 mm SL, Japan, Ryukyu Islands, Iriomote Island, coll. T. Suzuki, 22 August 1994. OMNH 39990, Japan, Ryukyu Islands, Iriomote Island, male, 28.5 mm SL, coll. T. Suzuki, 27 July 1997.
Additional material. CH 232 - 43 -006, 31.5 mm SL, Egypt, Dahab, coll. J. Herler, 15 June 2004; CH 232 - 43 - 0 15, juvenile, 19.2 mm SL, Egypt, Dahab, coll. J. Herler, 14 November 2005; CH 232 - 43 -040, 26.2 mm SL, Maldives, Fesdu Island, coll. J. Herler, 17 March 2007; PMR VP 3202, 32 mm SL, Egypt, Sharm el Sheikh, Sharm el Moya, coll. S.V. Bogorodsky, 0 4 July 2011; OMNH 39978, juvenile, 18.1 mm SL, Japan, Ryukyu Islands, Iriomote Island, coll. T. Suzuki, 0 8 August 1996. OMNH P- 40047 ( DNA sample), 30.3 mm SL, Japan, Ryukyu Islands, Iriomote Island, coll. T. Suzuki, M. Suzuki and A. Kawai, 22 August 2002.
Comparative material. Gobiodon unicolor ( Castelnau 1873) : holotype, MNHN A- 4015 ( Fig. 9 View FIGURE 9 I, J), 29.6 mm SL, Australia, Cape Sidmouth, coll. Castelnau.
Diagnosis. Dorsal-fin rays I, 10; anal-fin rays I, 8; head and body naked; obvious groove between isthmus and interopercle; body deep (the depth at pelvic-fin origin 40.4–44.3 % SL) and strongly compressed; head rounded in juveniles, large adults with steep profile, slightly hump-headed; caudal peduncle relatively slender (depth 14.8 – 16.0% SL). Juveniles and adults dark reddish brown with greenish subcutaneous gleam on dorsal part of body; median fins plain (western Pacific Oceans) or with pale margin (Red Sea and Indian Ocean); iris plain light blue (Red Sea) or scattered with red-brown dots on the outer margin.
Description (based on 5 types and several non-type specimens (for osteology)). Head and body strongly compressed. Body proportions and meristics for types are provided in Tables 8 and 9, respectively. Dorsal-fin rays VI + I, 10 (n = 8); anal-fin rays I, 8 (n = 8); pectoral-fin rays 19–20 (19:4, 20: 4); pelvic-fin rays I, 5, fin short (not reaching anus) and cup-shaped with significant frenum between spines; caudal fin with 15–17 segmented and branched rays. First dorsal fin lower than second dorsal and anal fins. Vertebral column with 10 precaudal and 16 caudal vertebrae, including urostyle. No scales. Gill opening as wide as pectoral-fin base, ending ventrally in opposite of 1 st or 2 nd lower pectoral-fin ray. Gill rakers 0–2 + 7–8. Obvious groove between interopercle and isthmus. Mouth slightly oblique, bending downwards and reaching approximately to below anterior margin of orbit. Upper lip usually slightly curved, slightly extending before snout. One outer row of 3 to 8 larger, slightly recurved teeth in upper and lower jaw, positioned on the anterior half of premaxilla and dentary. Medially, 3 to 4 rows of small, slender and recurved teeth in both jaws. In lower jaw, a pair of large, postsymphysial canines on each side, one or both on each side often small or absent, probably due to tooth loss and replacement. Anterior and posterior nasal openings at the end of short tubes. Head sensory canals typical as for Gobiodon ( Winterbottom & Harold 2005) , with anterior oculoscapular (pores NA (paired)), AI, PI (unpaired), SO, AO and IT (paired) and preopercular (three pores on each side) canals present.
Life colouration. Body uniformly dark reddish brown, occasionally with a greenish gleam on dorsal part of body (in Red Sea specimens), densely dotted with dark brown on body, nape, and pectoral-fin base ( Figs. 7 View FIGURE 7 A, C, 8); specimens from the Maldives characterised in having scattered tiny red spots; a weak purple streak of internal pigment runs along the lateral midline of the body, from behind the pectoral fins to the caudal-fin origin; iris plain light blue or scattered with red-brown dots on the outer margin; median fins with pale margin typically in specimens from the Red Sea and sometimes in specimens from the Maldives but fins plain in specimens from western Pacific Ocean.
Preserved colouration. Head and body uniformly dark brown. When mucous epidermis removed, body light brown with numerous small dark chromatophores scattered all over ( Fig. 7 View FIGURE 7 B). Iris bright. Pale margins on median fins often retained.
Molecular genetics. In the analysis here, the sequences of G.sp 3 _MA 1 from the Maldives and of G.sp 3 _GA 1 from the Gulf of Aqaba, Red Sea by Herler et al. (2009) are included. See Table 1 for Genbank accession numbers. Gobiodon fuscoruber sp. nov. is closely related to G. at er sp. nov. (see above). The intraspecific p -distances between populations from the Red Sea, the Maldives, Japan and the GBR (as “ G. unicolor ”) are around 0.02 (see Table 10), with the least distance between the two latter (0.001).
Habitat. On reef flats, crests and upper reef slopes in less exposed areas. Prefers Acropora selago but also occurs in a number of other, narrowly branched Acropora species such as A. acuminata and A. hyacinthus .
Distribution. The species is currently known from the Red Sea, central Indian Ocean and western Pacific Ocean ( Japan and GBR).
Etymology. This species is named after its uniformly reddish-brown life colouration. The name “ fuscoruber ” is a combination of the Latin words “ fuscus ” (= brown) and “ rubrum ” (= red). Although there are geographic colour variants (e.g. red dots in the Maldives), the reddish brown basic colouration is typical throughout its distribution area. Suggested common name: Brown-red Coralgoby.
western Pacific. Values are proportions of standard length (SL) and head length (last five measurements), respectively,
means and the first standard deviation (SD); d = damaged.
G. cit RS G. his RS G. r i v RS G. bil RS G. i r r RS G. ate RS G. f u s RS G. f u s Mal G. f u s Jap
G. cit GBR 0.031
G. h i s GBR 0.005
G. r i v GBR 0.023
G. b i l Mal 0.011
G. bil Tai 0.014
G. q u i Mal 0.033
G. re t RS 0.026
G. a t e Mal 0.016 G. a t e Tai 0.013 G. f u s RS 0.023 G. f u s Mal 0.026 0.019 G. f u s Jap 0.031 0.019 0.016 “ G. u n i ” GBR 0.032 0.020 0.017 0.001 Remarks. This species was informally designated as Gobiodon sp. 3 by Herler and Hilgers (2005). The Red Sea specimens are very similar to populations from the Maldives and the western Pacific, especially in body shape, meristics (D 2 I/ 10, A I/ 8 in all specimens from the Maldives and the western Pacific) and basic colour pattern. The Maldivian specimens, however, are somewhat brighter, the blue iris less obvious, and they have red dots on the postorbital area, nape, and body (more densely on the dorsum). They are similar to the Red Sea population in having reddish brown median fins, sometimes also with a pale margin. Specimens from Japan have a somewhat brighter body colouration (especially in juveniles) and the scattered dark brown chromatophores are more obvious than in the usually dark reddish brown Red Sea specimens. In addition, Japanese specimens often have two weak red bars below the orbit and several short red lines and dots on the anterior nape and snout. Suborbital bars may be retained in subadults but are very rarely seen in adults. Despite some geographic variation in colouration, the genetic distance of the populations investigated is very small. Therefore, we assume that these populations belong to one species.
Gobiodon fuscoruber was assumed to be identical with G. unicolor ( Castelnau 1873) after comparison with images in Munday et al. (1999) and Senou et al. (2004). Genetic investigations confirm that G. unicolor sensu Munday et al. (1999) and Harold et al. (2008) is identical with G. fuscoruber ( Fig. 11 View FIGURE 11 , Table 10). Examinations of the holotype of G. unicolor , however, revealed that this specimen represents a species with a black opercular spot (see Figs. 9 View FIGURE 9 I, J), which is characteristic for several species of the genus, but is present neither in G. fuscoruber nor in G. unicolor noted by any of the authors mentioned above. Considering the black opercular spot and the general body shape of the holotype of G. unicolor , the type species could be identical with G. histrio , G. axillaris or G. sp. C of Munday et al. (1999), which all have a black opercular spot. When comparing body shapes in detail, it becomes clear that the holotype represents G. histrio ( Valenciennes 1837) . This is confirmed by a comparison of the two preserved syntypes ( MNHN 3098) and x-ray images of G. histrio with the preserved holotype and x-ray image of G. unicolor : the types of both species resemble each other in that they have a deep and compressed body, a groove between the interopercle and isthmus, strongly re-curved jaws and identical fin ray counts in D 2 and A (10 and 9, respectively; except for one syntype of G. histrio that has 11 D 2 -rays). The colour description of G. histrio by Cuvier and Valenciennes (1837) and the range of fin rays in D 2 and A of the syntypes confirm its identity with G. histrio shown by recent authors (e.g., Munday et al. 1999, Herler & Hilgers 2005). Further evidence of the synonymy of G. histrio and G. unicolor comes from Castelnau’s (1873) description of the preserved colouration of G. unicolor as a light brown. When specimens of G. histrio are preserved, the typical green-red colouration fades very quickly after ethanol preservation and also turns uniformly light brown (see Herler & Hilgers 2005).
The final confirmation of the synonymy of G. unicolor and G. histrio comes from a geometric morphometric analysis. Principal component analysis plotted the holotype of G. unicolor clearly within the morphospace of G. histrio ( Fig. 10 View FIGURE 10 ), and discriminant function analysis on the scores of the first 6 PCs of all 111 specimens revealed a 99.99 % probability of assignment to G. histrio . We therefore propose G. unicolor ( Castelnau 1873) to be a junior synonym of G. histrio ( Valenciennes 1837) and as not applicable to the uniformly coloured species described herein. The new name G. fuscoruber not only has to be used for the species called G. unicolor by several authors mentioned above, but also applies to some undescribed species, such as G. sp. 5 of Akihito et al. (2002), and G. sp. D of Senou et al. (2004). Apart from genetics (at least tested for “ G. unicolor ”), the uniformly (brownish) life colour, the rounded head shape, the low first dorsal fin and the large pectoral fins unite these taxa. This suggests that G. fuscoruber is a widely distributed species, occurring throughout the Indo-Pacific reef province.
Gobiodon fuscoruber and G. ater share a special feature with at least seven other species of the genus: a groove between the interopercle and isthmus ( Harold & Winterbottom 1999; Harold et al. 2008), which is also present in G. brochus Harold and Winterbottom 1999 and G. flavus Sauvage 1880 . However, these species are clearly distinct from G. at e r and G. fuscoruber in that they have a much brighter colouration. Furthermore, G. f l a v us and G. brochus have at least 9 anal fin rays ( Sauvage 1880, Harold & Winterbottom 1999) instead of the very constant number of 8 rays in G. fuscoruber and G. at e r. Moreover, the colour description of Sauvage (1880) for G. f l av u s notes a yellowish colouration. The combination of this colouration and presence of the groove makes assignment of G. flavus to any of the currently known species difficult, though it is reasonable to conclude that it is not identical with any of the species described herein. Other species with such a groove, like G. sp. A, sp. B, sp. C, G. histrio and G. erythrospilus ( Harold et al. 2008) are very different in their life coloration and cannot be mistaken with G. fuscoruber or G. at e r.




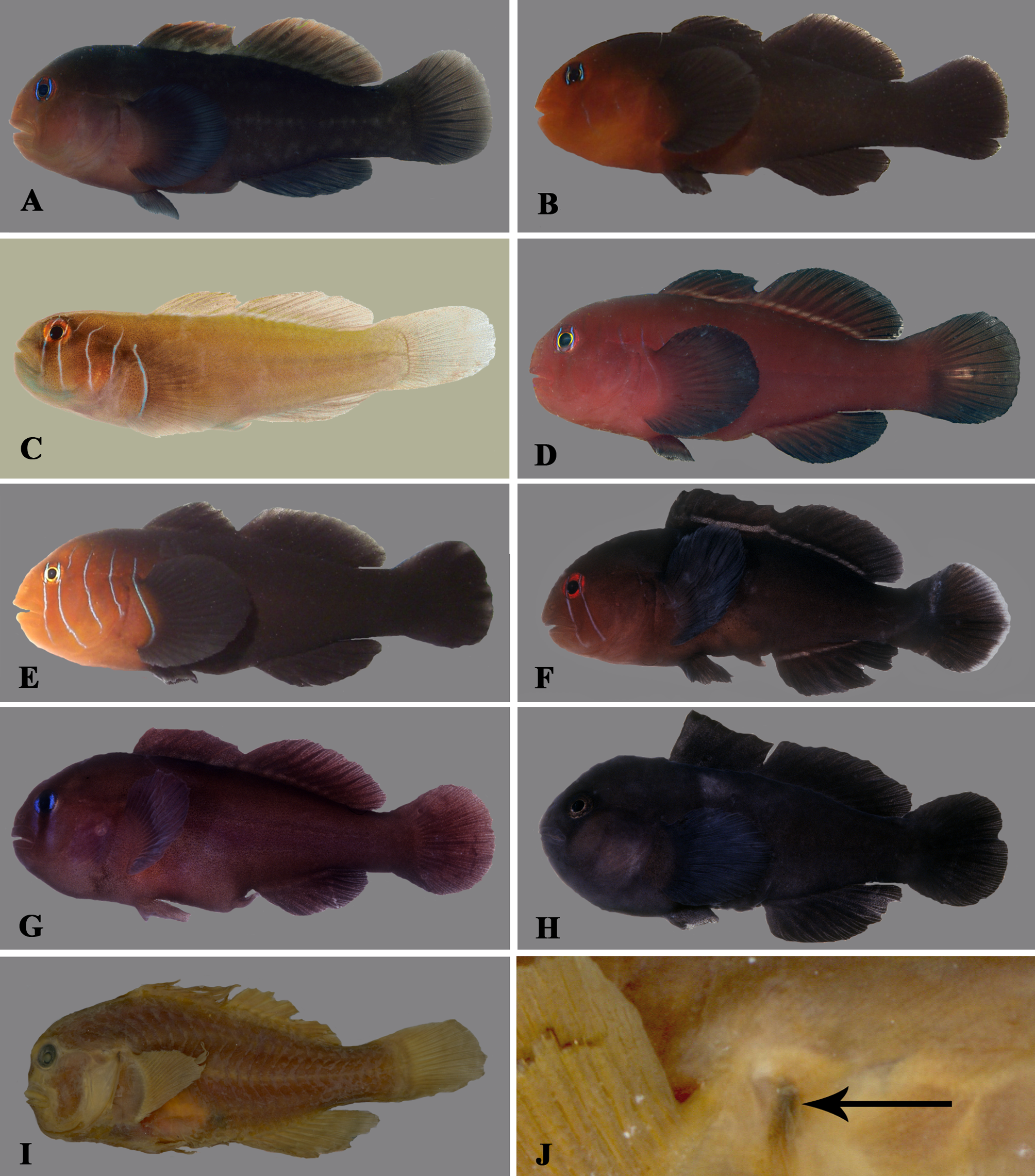

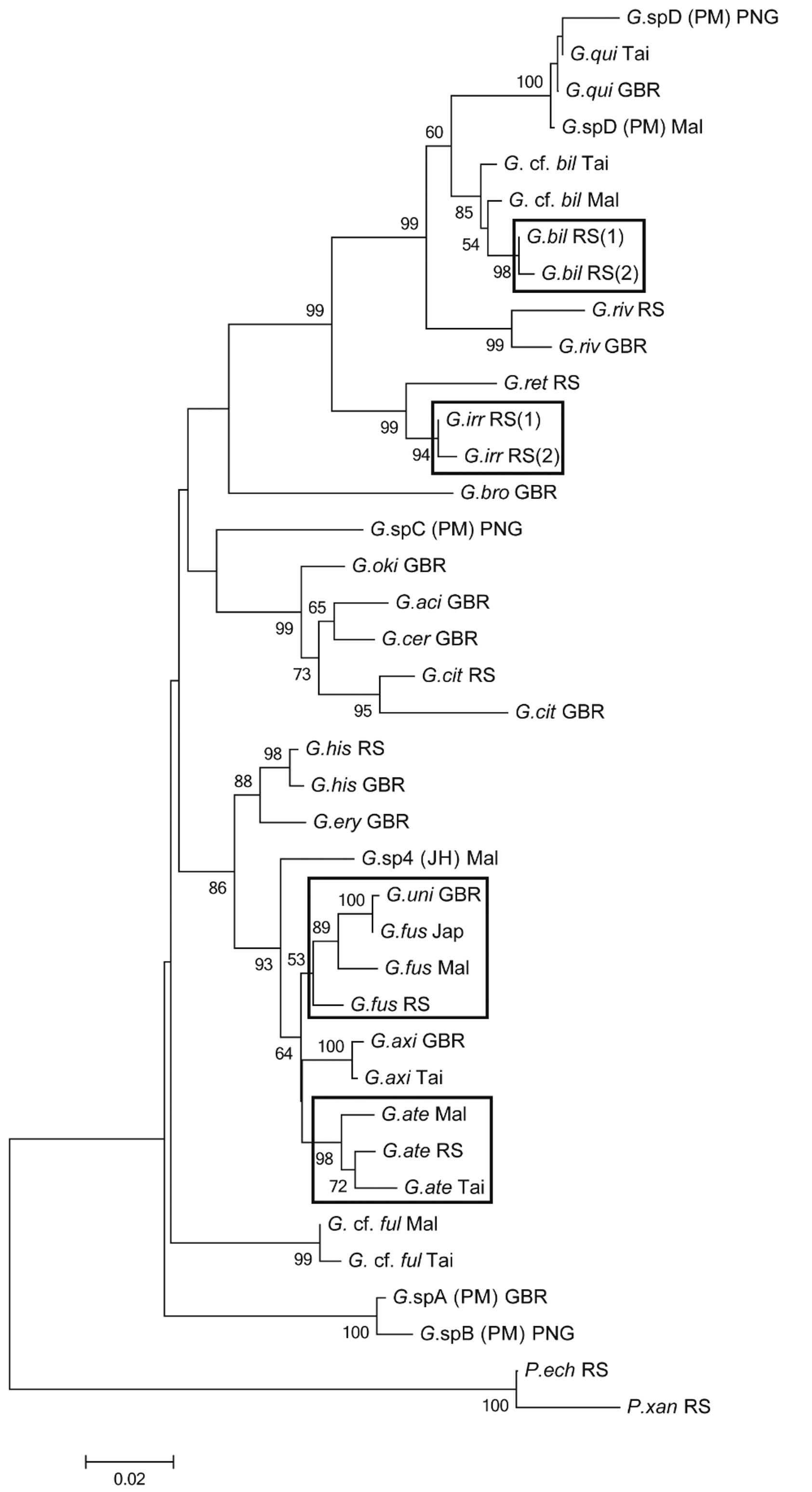
TABLE 8. Body proportions of holotype and seven paratypes of Gobiodon fuscoruber sp. nov. from the Red Sea and
| Status Coll.No. | Holo-type NMW 95079 | Para-type NMW 95080 | Para-type MNHN 2006- 1700 | Para-type BMNH 1951-1- 16-554 | Para-type BMNH 1951-1- 16-555 | Para-type OMNH 39984 | Para-type OMNH 39986 | Para-type OMNH 39990 | MEAN (±SD) |
| SL (mm) | 36.7 | 31.3 | 32.1 | 29.2 | 34.0 | 29.2 | 23.6 | 28.5 | |
| Snout to first dorsal-fin origin | 39.8 | 41.1 | 42.6 | 40.3 | 38.2 | 37.4 | 36.5 | 38.2 | 39.3 (2.0) |
| First dorsal-fin origin to second dorsal-fin origin | 25.5 | 24.5 | 25.2 | 21.9 | 21.9 | 21.3 | 22.1 | 23.2 | 23.2 (1.6) |
| Second dorsal-fin origin to anal-fin origin | 36.9 | 35.2 | 37.2 | 35.0 | 33.6 | 32.9 | 32.2 | 33.1 | 34.5 (1.9) |
| Pelvic-fin origin to anal-fin origin | 25.2 | 25.0 | 25.4 | 29.8 | 29.1 | 28.2 | 28.5 | 26.5 | 27.2 (1.9) |
| Snout to pelvic-fin origin | 39.9 | 42.0 | 40.9 | 38.1 | 39.1 | 38.0 | 38.1 | 41.6 | 39.7 (1.6) |
| First dorsal-fin origin to pelvic-fin origin | 42.7 | 42.1 | 44.3 | 41.2 | 40.4 | 39.0 | 38.5 | 41.4 | 41.2 (1.9) |
| First dorsal-fin origin to anal-fin origin | 48.5 | 47.5 | 47.7 | 47.6 | 46.0 | 42.9 | 44.4 | 45.1 | 46.2 (2.0) |
| Pelvic-fin origin to second dorsal-fin origin | 45.3 | 43.9 | 46.0 | 44.1 | 43.4 | 43.4 | 42.4 | 43.5 | 44.0 (1.1) |
| Head length | 29.6 | 32.4 | 32.0 | 31.1 | 29.1 | 28.1 | 28.3 | 29.3 | 30.0 (1.7) |
| Head depth | 35.5 | 33.8 | 36.2 | 34.0 | 35.1 | 35.4 | 34.4 | 38.4 | 35.3 (1.5) |
| Body depth | 42.9 | 40.9 | 43.8 | 40.9 | 40.1 | 39.6 | 38.3 | 41.2 | 41.0 (1.7) |
| Pelvic-fin length | 16.1 | 16.4 | 18.3 | 16.6 | 17.0 | 16.1 | 16.4 | 15.4 | 16.5 (0.8) |
| Anal-fin length | 21.1 | 20.3 | 20.5 | 19.3 | 20.1 | 21.2 | 21.1 | 21.9 | 20.7 (0.8) |
| Second dorsal-fin length | 27.9 | 28.8 | 30.1 | 27.8 | 28.7 | 31.5 | 31.3 | 28.1 | 29.3 (1.5) |
| Caudal fin length | 23.4 | 24.6 | 23.7 | 23.8 | d | 22.4 | 19.1 | 20.4 | 22.5 (2.0) |
| Pectoral-fin length | 26.6 | 25.4 | 26.2 | d | d | 26.4 | 26.9 | 24.2 | 26.0 (1.0) |
| Caudal-peduncle length | 21.4 | 22.2 | 21.9 | 22.9 | 23.7 | 21.6 | 22.5 | 21.2 | 22.2 (0.8) |
| Caudal-peduncle depth | 15.7 | 14.8 | 15.4 | 16.0 | 15.2 | 14.1 | 15.6 | 15.4 | 15.3 (0.6) |
| Interorbital width | 18.5 | 14.9 | 16.4 | 17.0 | 17.6 | 17.4 | 18.6 | 17.1 | 17.2 (1.2) |
| Horizontal-eye diameter | 20.2 | 18.1 | 19.0 | 18.9 | 19.0 | 19.9 | 21.1 | 19.9 | 19.5 (0.9) |
| Snout length | 29.7 | 31.9 | 33.9 | 29.8 | 30.1 | 27.3 | 26.2 | 26.0 | 29.4 (2.8) |
| Upper-jaw length | 36.0 | 32.0 | 36.7 | 29.8 | 30.7 | 28.5 | 28.0 | 31.8 | 31.7 (3.2) |
| First dorsal spine to first pectoral ray | 69.2 | 62.1 | 62.7 | 59.8 | 63.3 | 50.1 | 50.5 | 55.5 | 59.2 (6.6) |
TABLE 10. Selection of genetic (p -) distances from pairwise comparisons of specimens of G. bilineatus sp. nov., G. irregularis sp. nov., G. ater sp. nov. and G. fuscoruber sp. nov. from different regions (GBR: Great Barrier Reef, Jap: Japan, Mal: Maldives, RS: Red Sea, Tai: Taiwan). For comparison of intra- and interspecific genetic distances across the genus, other species from different regions as well as “ G. unicolor ” (sensu Munday et al. (1999) and Harold et al. (2008 )) from the GBR are included. Italic numbers indicate intraspecific distances. Bold values indicate intraspecific distances of more than 2 %. For species abbreviations and Genbank accession numbers see Table 1.
| Status Holotype Coll.No. NMW 95079 | Paratype NMW 95080 | Paratype Paratype Paratype Paratype MNHN BMNH 1951- BMNH 1951- OMNH 2006-1700 1-16-554 1-16-555 39984 | Paratype OMNH 39986 | Paratype OMNH 39990 |
| D1 VI | VI | VI VI VI VI | VI | VI |
| D2 10 | 10 | 10 10 10 10 | 10 | 10 |
| A 8 | 8 | 8 8 8 8 | 8 | 8 |
| C (br.) 17 | 17 | 17 17 17 17 | 17 | 17 |
| C (segm.) 17 | 17 | 17 17 17 17 | 17 | 17 |
| P 20 | 20 | 20 19 20 19 | 19 | 19 |
| V I/5+I/5 | I/5+I/5 | I/5+I/5 I/5+I/5 I/5+I/5 I/5+I/5 | I/5+I/5 | I/5+I/5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gobiodon fuscoruber
| Herler, Juergen, Bogorodsky, Sergey V. & Suzuki, Toshiyuki 2013 |
Gobiodon
| Herler 2005: 123 |
| Herler et al. (2009) : 733 |
Gobiodon unicolor
| Hayashi 2003: 63 |
| Munday et al. (1999) : 56 |
| Senou et al. (2004) : 170 |
| Harold et al. (2008) : 132 |
Gobiodon
| Senou et al. (2004) : 175 |