
Chusquea calderoniae K.V.A.Vidal & L.G.Clark, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.579.4.3 |
|
DOI |
https://doi.org/10.5281/zenodo.7568669 |
|
persistent identifier |
https://treatment.plazi.org/id/03E07458-CF6E-D654-F598-C83FFCDA141F |
|
treatment provided by |
Plazi |
|
scientific name |
Chusquea calderoniae K.V.A.Vidal & L.G.Clark |
| status |
sp. nov. |
Chusquea calderoniae K.V.A.Vidal & L.G.Clark sp. nov.
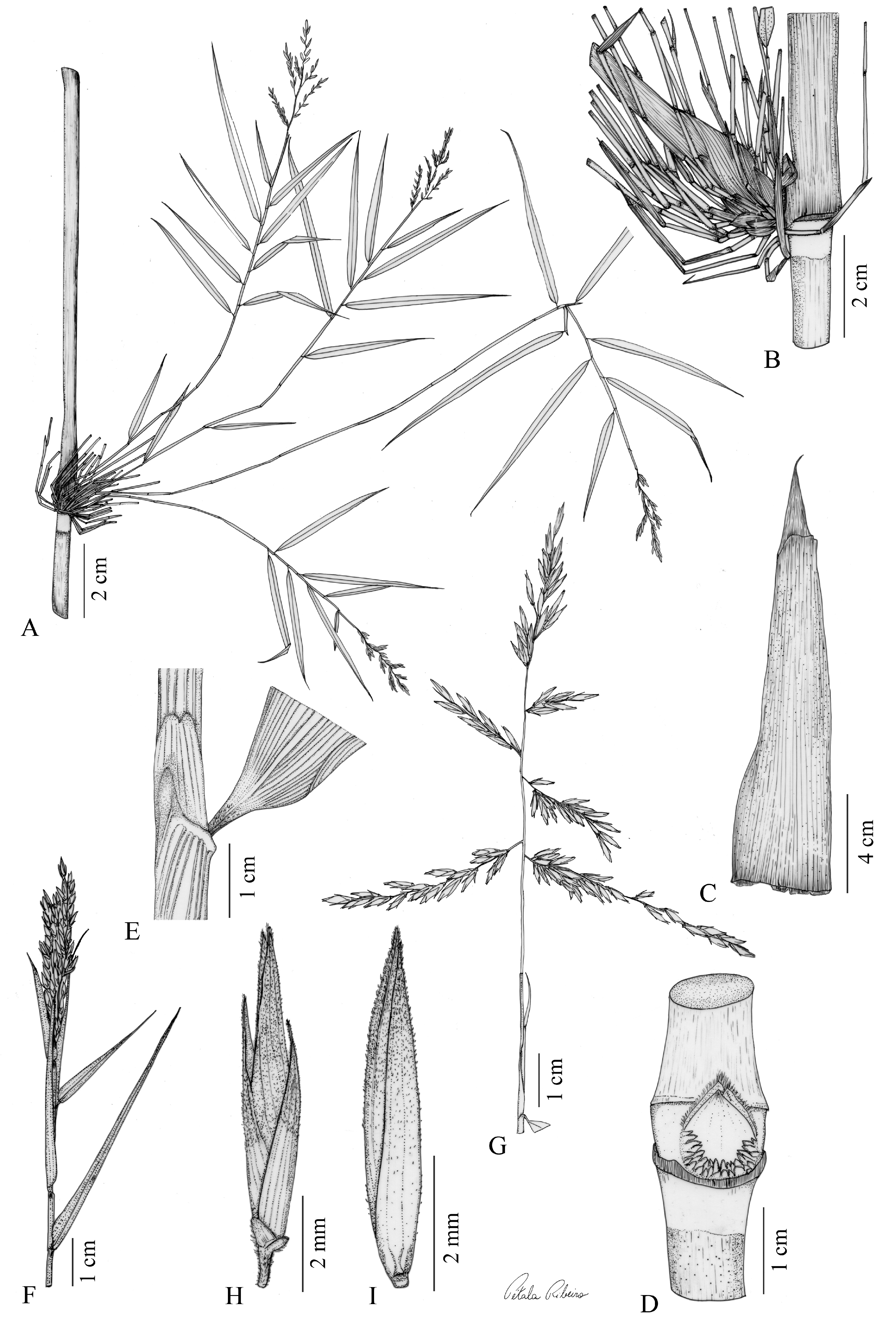
TYPE:— BRAZIL. Bahia, Mun. Rio de Contas, fragmento de Mata Atlântica próximo à base do Pico das Almas , regi ã o de córrego do rio, nascente do Rio Brumado , ca. 3.5 km de distância da estrada principal (text in Portuguese), 13°31’29’’S, 41°57’24’’W, 1489 m, 13 June 2016 (fl.), Vidal & Clark 241 (holotype HUEFS!, isotypes CEPEC!, ISC!, SP!, SPF!, US!). Figs. 1 View FIGURE 1 , 2 View FIGURE 2 GoogleMaps .
Diagnosis:— Chusquea calderoniae differs from C. attenuata mainly by the presence of a white waxy infranodal band (vs. absent), culm leaves with blades triangular to narrow-triangular (vs. triangular to deltate), central bud triangular in outline (vs. circular), longer subsidiary branches [(20.5‒) 31‒45 (‒68.5) cm long vs. (9.1‒) 24‒33.5 cm], as well as foliage leaf sheaths with a longer inner ligule on the underlapping margin [2.5‒9 mm long vs. (0.4‒) 0.5‒1.7 mm]. Reproductively, the new species bears shorter spikelets [5‒6 mm long vs. (5.1‒) 7.1‒8.5 mm] and shorter glumes III [2.7‒3.2 mm long vs. (3.5‒) 5‒6 mm] and IV [3.1‒4 mm long vs. (3.7‒) 4.8‒7 mm], as well as shorter lemmas [4‒5.1 mm long vs. (5.2‒) 6.1‒7.8 mm] and paleas (4.4‒5.2 mm long vs. 5‒6.6 mm).
Description: —Rhizomes unknown. Culms 4‒5 m tall, (0.65‒) 0.9‒1.2 cm in diameter, erect at the base then arching or scandent towards the apex; internodes 21.5‒25 cm long, terete, solid, green to vinaceous, glabrous except scabrous with a white waxy band (2‒3 cm long) just below the node. Culm leaves (7.2‒) 8.4‒12.4 cm long, erect, not reaching the next node, the juncture of the sheath and blade abaxially a faint line or obscure; sheaths (6‒) 6.5‒9.8 cm long, (0.8‒) 1.4‒1.8 cm wide, 2.3‒6 (‒12) times as long as the blade, persistent, linear-triangular, margins glabrous, the overlapping one fused to the sheath at the base for (0.2‒) 0.5‒1 cm, abaxially scabrous to slightly scabrous towards the apex, adaxially glabrous and shiny, tessellate; summit extensions absent; girdles (2.8‒) 7‒9 mm long, stramineous to brown, pilose, no prominent skirt but with a slight corky ridge at the juncture with the sheath; outer ligule absent; inner ligule 0.2‒0.5 (‒1) mm long, erect, rigid or sometimes slightly membranous, stramineous to brown, glabrous or less frequently pubescent, apex truncate and long ciliate; blades (0.6‒) 1.2‒2.8 (‒3.5) cm long, 0.3‒0.5 cm wide, triangular to narrow-triangular, non-pseudopetiolate, erect, persistent, abaxially scabrous, adaxially long-pilose to scabrous, slightly tessellate, the midrib evident only at the tip, margins glabrous, apex long setose, 0.3‒0.7 mm long. Nodes at mid-culm with a triangular central bud subtended by 18‒36 smaller, subequal subsidiary buds in 2‒3 rows, in a constellate arrangement; central bud prophyll glabrous with margins long ciliate; nodal line dipping slightly below the bud/branch complement; nodal region 0.5‒0.7 mm long; supranodal ridge visible, slightly raised and prominent; nodal line raised and prominent. Branching infra-extravaginal, being initially infravaginal, often becoming also extravaginal as branches develop, leafy subsidiary branches 18‒36 per node, (20.5‒) 31‒45 (‒68.5) cm long, subequal, non-rebranching, non-geniculate, arching and ascending. Foliage leaves 6‒11 per complement; sheaths 2.9‒4.5 cm long, glabrous to scabrous or hispid at the apex, stramineous, keeled on the upper half, the margins long ciliate on only one side, summit extension on both sides, being fused to the inner ligule on underlapping one, 0.5‒1.2 mm long, subequal, scabrous to hispid; outer ligule asymmetrical and 2-lobed, scabrous (less frequently glabrous), erect and rigid, apically ciliolate, the smaller side 0.2‒0.4 mm long, the bigger one 0.35‒0.8 mm long; inner ligule asymmetrical, membranous, scabrous, glabrous at the apex, towards the overlapping margin, 0.2‒0.4 mm long, truncate and free from the summit extension, on the underlapping one 2.5‒9 mm long, attenuate and fused to the summit extension; pseudopetioles 1‒3 mm long, thick, distinct to somewhat distinct, abaxially glabrescent with widely spaced trichomes, adaxially pilose with deposition of wax, pulvinus brown when present; blades 4.1‒12 cm long, 0.3‒0.8 cm wide, L:W 13.5‒27.6, linear-lanceolate, non-tessellate, abaxially scabrous and rarely with a tuft of trichomes at the base, adaxially pilose, sometimes scabrous only at the base, base attenuate, midrib slightly excentric, abaxially prominent throughout the blade except at the apex, adaxially prominent only on the lower half of the blade, both margins scabrous, apex long setose. Synflorescences (4.5‒) 5‒9.4 cm long, (3‒) 3.6‒8 cm in diameter, paniculate, open, pyramidal, with the lower branches slightly reflexed to reflexed, subtended by 1‒2 spatheate bracts, these slightly scabrous at the base becoming scabrous-pilose towards the apex, the first (lower) one less differentiated, its sheath (2.6‒) 3.8‒7.1 cm long, papyraceous, stramineous to greenish, a band of trichomes around the node, its blade 1.6‒2.1 cm long, green, linearlanceolate, apex long setose, the second (upper) one differentiated, its sheath (3.7‒) 7.4‒9.9 cm long, papyraceous, its blade 0.4‒1.1 cm long, green to stramineous, linear-lanceolate with apex long setose; rachis (3.9‒) 7.5‒10.5 mm long, triquetrous, densely pilose; branches and pedicels angular, densely pilose to hispid, all subtended by a scar or rim or occasionally a scale-like subtending bract, the 1‒3 lowermost branches erect when young and divergent (horizontal) to reflexed at maturity, the lowermost one 1‒4.5 cm long, the second 1.2‒4.3 cm long, and the third 1.2‒4.3 cm long, the next higher 1‒3 branches (fourth through sixth) ascending when young and slightly reflexed at maturity, the fourth (1‒) 2‒2.8 cm long, the fifth (1‒) 1.5‒2.2 cm long, and the sixth 1‒2 cm long, the next higher 1‒3 middleupper branches (seventh through ninth) erect when young and reflexed to strongly reflexed at maturity, the seventh 0.8‒1.5 cm long, the eighth 0.7‒1 cm long, and ninth ca. 1 cm long, the tenth and higher branches and pedicels erect to appressed to the primary branches; pedicels 0.5‒1.3 (‒2.8) mm long. Spikelets 5‒6 mm long, 0.9‒1.2 mm wide, dorsally compressed; glumes I and II scale-like, obtuse, non-nerved; glume I 0.2‒0.35 mm long, <1/25 the spikelet length, slightly pubescent at the base, becoming glabrous towards the apex; glume II 0.2‒0.45 mm long, <1/20 the spikelet length, glabrous; glumes III and IV lanceolate, slightly navicular for the upper half, non-keeled, abaxially slightly glabrescent to pubescent at the base then pilose towards the apex, 3-nerved; glume III 2.7‒3.2 mm long, ca. 1/2 the spikelet length, apex mucronate to apiculate (the apicule 0.1‒0.35 mm long), margins ciliate; glume IV 3.1‒4 mm long, 3/5‒2/3 the spikelet length, apex aristulate (the aristule 0.15‒0.25 mm long); lemma 4‒5.1 mm long, somewhat navicular on the upper half, keeled, apex acute, awned (the awn 0.4‒0.5 mm long), hyaline to greenish-hyaline (shiny), abaxially glabrescent at the base, becoming pubescent with a tuft of trichomes at the apex, 5‒7-nerved; palea 4.4‒5.2 mm long, 2-apiculate (the apicules 0.1‒0.3 mm long), 4-nerved, 2-keeled, strongly sulcate only for the upper half, the sulcus scabrid, abaxially pubescent then pilose towards the apex. Lodicules 3, glabrous, hyaline, apically long ciliate, 1-nerved; anterior pair 0.8‒1 mm long, ovate, the posterior one 0.4‒0.6 mm long, lanceolate. Stamens 3; anthers 1.6‒2 mm long, brownish to whitish-brown. Caryopsis not seen.
Etymology: —The specific epithet honors the Argentinian bamboo researcher Cleofé E. Calderón (1929‒2007), who first collected this species, and who made great contributions to research in bamboos and other grasses worldwide ( Clark et al. 2008).
Taxonomic notes: —According to the current classification ( Clark 2004, Fisher et al. 2014, Andrade et al. 2019, Vidal et al. 2021, Clark et al. 2022), eight species are recognized in the C. meyeriana informal group: C. anelythra, C. anelytroides Ruprecht ex D̂ll (1880: 206), C. attenuata , C. clemirae Mota, Oliveira & Clark in Mota et al. (2013: 95), C. cordata, C. longispiculata Clark (2004: 34), C. meyeriana, and C. parviligulata Andrade, Pianissola & Clark in Andrade et al. (2019: 29), all of them endemic to Brazil, occurring in the Atlantic rainforest from the state of Rio Grande do Sul to Bahia ( Clark et al. 2020, Vidal et al. 2021, Clark et al. 2022). This group is recognized by several morphological similarities, such as the presence of a white waxy infranodal band, culm leaves with at least a slight corky ridge at the juncture of the girdle and sheath, at least initially infravaginal branching, synflorescence branches subtended by 1‒4 spatheate bracts, at least the lower two primary synflorescence branches strongly reflexed, and spikelets with very reduced glumes I and II ( Clark 2004, Andrade et al. 2019, Vidal et al. 2021, Clark et al. 2022).
Chusquea calderoniae is herein included in the C. meyeriana informal group, due to its clear morphological similarities to C. attenuata , considering that samples were previously misidentified as “ C. attenuata ” or annotated as “ C. aff. attenuata ” in herbarium collections. In fact, these two taxa share several vegetative characters ( Table 1 View TABLE 1 ), such as culm leaves not reaching the next node, with blades of similar length and not very well-differentiated from the sheaths, in addition to subsidiary buds arranged in 1‒3 rows, foliage leaves with similar-sized outer ligules, inner ligules asymmetrical (but these can also be attenuate in C. attenuata ), and blades linear-lanceolate. Reproductively, these species can be recognized by the presence of up to 1‒2 spatheate bracts subtending the synflorescences, and spikelets bearing glumes III and IV not reaching the full spikelet length, even though they differ in lengths ( Table 1 View TABLE 1 , Fig. 1 View FIGURE 1 ).
However, despite these similarities, Chusquea calderoniae exhibits several morphological differences from C. attenuata ( Table 1 View TABLE 1 ). Vegetatively, it can be recognized by the presence of a white waxy infranodal band 2‒3 cm long (vs. absent); shorter culm leaves [(7.2‒) 8.4‒12.4 cm long vs. 12.4‒16.5 cm], with blades triangular to narrowtriangular (vs. triangular to deltate), in addition to the sheaths shorter [(6‒) 6.5‒9.8 cm long vs. 11‒14.7 cm]; subsidiary branches arching and ascending (vs. straight, geniculate to slightly geniculate at the basal nodes) and longer [(20.5‒) 31‒45 (‒68.5) cm long vs. (9.1‒) 24‒33.5 cm]; foliage leaves with the outer ligule asymmetrical and 2-lobed [vs. U-shaped, less usually asymmetrical, one side truncate (smaller) and other 1-lobed (bigger)], and longer inner ligule on the underlapping margin [2.5‒9 mm long vs. (0.4‒) 0.5‒1.7 mm]; and blades abaxially scabrous and rarely with a tuft of trichomes at the base (vs. pilose, with a tuft of trichomes at the base) and adaxially pilose, sometimes scabrous only at the base (vs. scabrous to hispid-scabrous, sometimes slightly scabrous on the upper half). Reproductively, C. calderoniae presents smaller spikelets [5‒6 mm long vs. (5.1‒) 7.1‒8.5 mm], as well as shorter spikelet bracts: glume III [2.7‒3.2 mm long, ca. 1/2 the length of the spikelet vs. (3.5‒) 5‒6 mm, 3/4 to 4/5 of it], glume IV [3.1‒4 mm long, 3/5 to 2/3 the length of the spikelet vs. (3.7‒) 4.8‒7 mm, 4/5 to 5/6 of it], lemma [4‒5.1 mm long vs. (5.2‒) 6.1‒7.8 mm], and palea (4.4‒5.2 mm long vs. 5‒6.6 mm). Additionally, the new species can also be differentiated by having glume III mucronate to apiculate (vs. awned), glume IV aristulate (vs. awned), and the palea 2-apiculate (vs. 2-awned, less usually 2-apiculate).
Characteristics of the culm leaves and the central and subsidiary buds are important features in the taxonomy of Chusquea ( Clark 1989, 1992, 1993, 1997, Vidal et al. 2018, 2021, Andrade et al. 2019, Ruiz-Sanchez et al. 2021 b, 2022, Clark et al. 2022, McMurchie et al. 2022). We observed two main morphological groups within the C. meyeriana group based on the shape of the central bud: circular and triangular.Although C. calderoniae shares a triangular central bud with C. clemirae, C. cordata, C. longispiculata, and parviligulata, the new species differs from those four by having culm leaves not reaching the next node (vs. reaching or surpassing the next node), and with the blade nondifferentiated (vs. blades easily differentiated). On the other hand, these characters bring C. calderoniae closer to C. anelythra, C. anelytroides, C. attenuata , and C. meyeriana, which all have circular central buds.
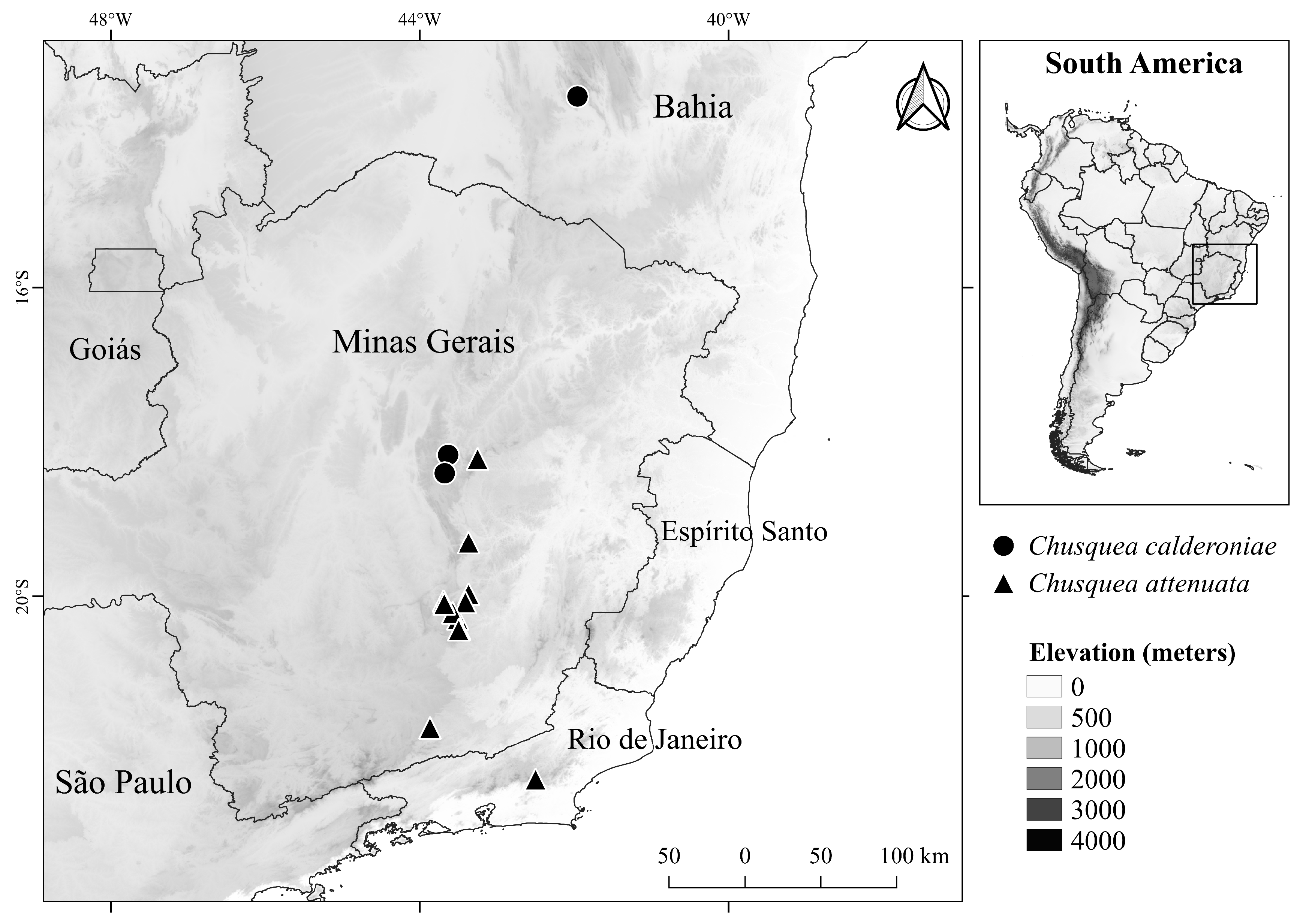
Distribution and habitat: — Chusquea calderoniae is currently known from only three populations distributed throughout the Espinhaço Range, one from Pico das Almas, which is part of the Chapada Diamantina province, in the state of Bahia, and the other two from the Diamantina Plateau district, located in the Southern Espinhaço province, in the state of Minas Gerais ( Colli-Silva et al. 2019) ( Fig. 3 View FIGURE 3 ). These specimens were collected in forest fragments, at 1100–1500 m.a.s.l, and always around rivers. The flora where this species occurs is usually rich in trees, with a partially closed canopy and herbaceous substrate, occurring together with representatives of Fabaceae , Anacardiaceae , and ferns.
Analysis of the herbarium samples of Chusquea attenuata indicates the occurrence of this species in Atlantic rainforest fragments of eastern Brazil, mostly in Meridional Espinhaço Range towards campos de altitude, from the eastern portion of Minas Gerais to Rio de Janeiro states ( Campos et al. 2017, Vidal et al. unpubl. data). Although both C. calderoniae and C. attenuata occur towards the southern limit of the Espinhaço Range (municipalities of Ouro Branco and Ouro Preto in Minas Gerais state), they occupy different bioregions and phytophysiognomies in this region ( Colli-Silva et al. 2019). The new species is restricted to the Espinhaço Range, mainly in forest fragments associated with campo rupestre vegetation, whereas C. attenuata occurs in the Atlantic rainforest domain, being restricted to southeastern Brazil, and generally associated with forests or campos de altitude from Ouro Preto (Minas Gerais) to the state of Rio de Janeiro ( Clark et al. 2020).
Conservation status: —The estimated area of occupancy (AOO) of Chusquea calderoniae is 12 km ², whereas the extent of occurrence (EOO) is 1,183 km ². Following the IUCN Red List categories and criteria ( IUCN Standards and Petitions Committee 2022), this new species should be considered Endangered (EN) [B1, B2ab(iii)] since it has a reduced number of documented populations (only three) and an AOO less than 500 km ² and an EOO less than 5,000 km ², in addition to being restricted to forest fragments associated with campo rupestre vegetation, whose areas are under intensely anthropic actions, even in environmental protection areas ( Mota et al. 2013, Clark et al. 2022). According to CNCFlora (2022), C. attenuata is also considered Endangered (EN) [A2c] ( Clark et al. 2020); however, the circumscription of this species is under study to provide accurate information about its conservation status (Vidal et al. unpubl. data).
Phenology: —Two flowering collections of Chusquea calderoniae from the state of Bahia were found, collected in June 2016 (Vidal & Clark 241) and April 2017 (Pianissola & Clark 174). The collections from the state of Minas Gerais were in the vegetative stage: one in February 1990 (Clark & Morel 711) and another in April 2016 (Vidal et al. 210, 211, 212), the latter in the same year as the flowering one from Bahia state (Vidal & Clark 241). Even in the vegetative stage, the characters were enough to confirm their identification. These data make it difficult to accurately infer anything about the flowering cycle of this new species. Thus, we cannot discard the possibility of an asynchronous or sporadic flowering event in this species. In any case, a flowering event extending for up to two years may be inferred.
Additional specimens examined (paratypes): — BRAZIL. Bahia: municipality of Rio de Contas, Pico das Almas , “Campo do Queiros” 19 km by car of Rio de Contas and 4 km walking, 1400 m, 11 April 1977 (veg.), Calderón 2458 (SP, US) GoogleMaps ; idem, 10 August 2000 (veg.), Oliveira 606 (HUEFS) GoogleMaps ; idem, 22 January 2017 (veg.), Mascarenhas 315 (HUEFS) GoogleMaps ; idem, 13°31’29’’S, 41°57’24’’W, 1489 m, 19 April 2017 (fl.), Pianissola & Clark 174 (HUEFS) GoogleMaps . Minas Gerais: municipality of Diamantina, road west from Diamantina to Biribiri, on the other side of Riber ã o das Pedras about 0.5 km from Biribiri, 18°10’S 43°38’W, 1100 m, 27 February 1990 (veg.), Clark & Morel 711 (BHCB, ISC, RB, SJR, SP, US) GoogleMaps ; same municipality, estrada Diamantina/Datas, caminho a esquerda antes da placa de limite de município Diamantina-Datas, cerca de 1 km dentro da trilha, córrego debaixo de ponte em uma bifurcaç ã o, pegando o caminho a esquerda, 19 April 2016 (veg.), Vidal et al. 210, 211, 212 (HUEFS) GoogleMaps .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |