
Axinella spatula, Sitjà & Maldonado, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3760.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:E05CF7B1-8410-4482-AB7D-DC9833479CC3 |
|
DOI |
https://doi.org/10.5281/zenodo.5042239 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF87B6-2977-FFCE-FF30-F948C968FA5E |
|
treatment provided by |
Felipe |
|
scientific name |
Axinella spatula |
| status |
sp. nov. |
Axinella spatula nov. sp.
( Figs. 6D–G View FIGURE 6 , 8 View FIGURE 8 ; tables 2, 3, 5)
Etymology. The species is named after the "spatula" tool (a diminutive form of the Latin "spatha"), which bears resemblance to the external shape of the specimens.
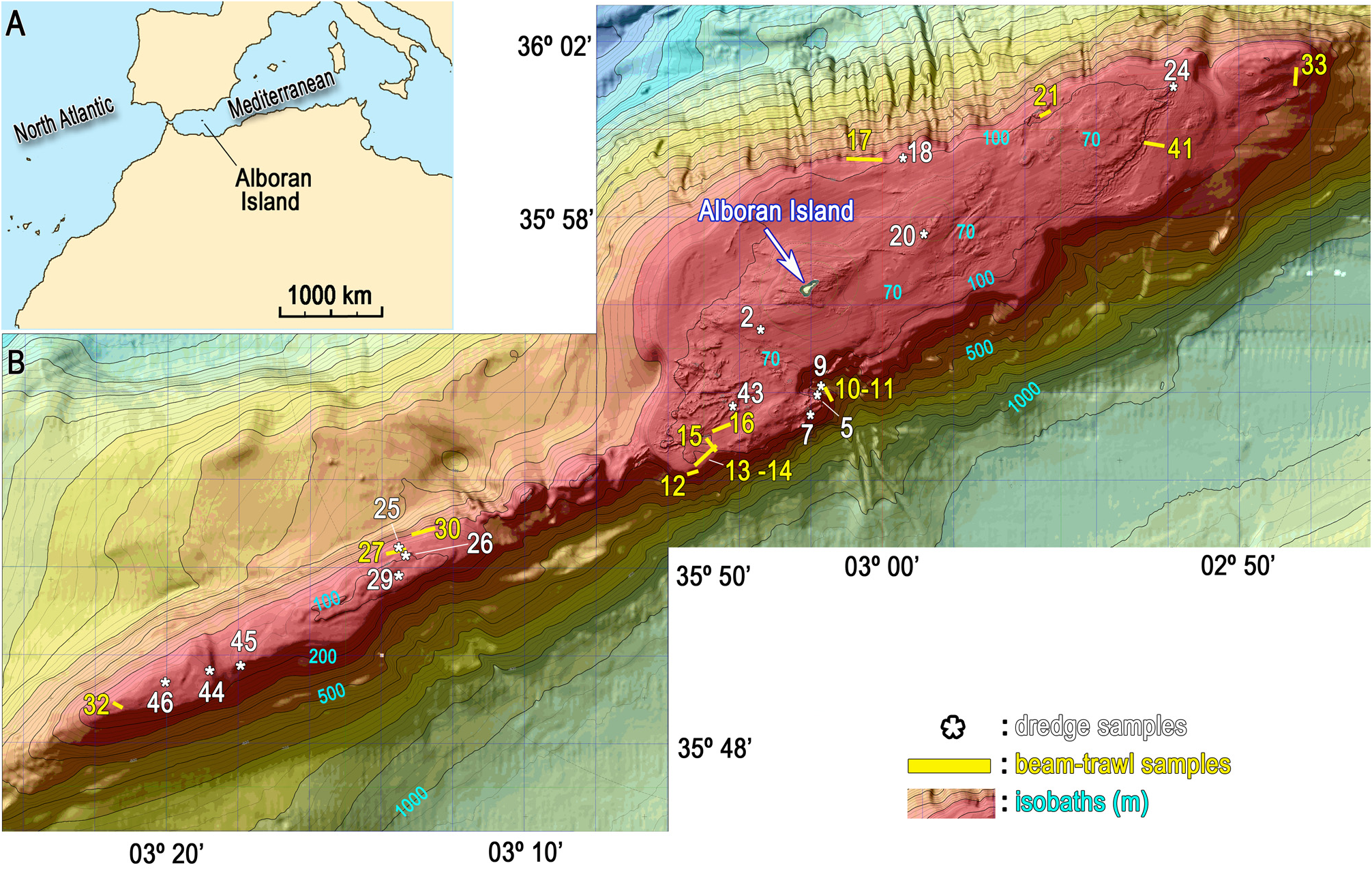
Material examined. Holotype MNCN-Sp145-BV33B collected from type locality Stn. 33 ( Table 1 View TABLE 1 , Fig. 1 View FIGURE 1 ), a 134 to 173 m deep, gravel bottom at the deep shelf of the Alboran Island.
Twenty-two paratypes designated: MNCN-Sp28-BV14A to C from Stn. 14; MNCN-Sp116-BV15A to I from Stn.15; MNCN-Sp57-BV21A to C (blackish specimens) and MNCN-Sp65-BV21A & B from Stn. 21; MNCN- Sp145-BV33A & C to D from Stn. 33; MNCN-Sp188-BV41A & B from Stn. 41 ( Table 1 View TABLE 1 , Fig. 1 View FIGURE 1 ).
Comparative material: Syntype material of Axinella flustra ( Topsent, 1892) = Tragosia flustra Topsent, 1892 , since no holotype was designated by Topsent (1892) for this species (Table 3). Syntypes were two specimens (MOM-040044) from Bay of Biscay (Stn. 58; 43º40’N 8º55’W, 134 m deep, 7 August 1886) and two specimens (MOM-040272) from Azores (Stn. 247; 38º23.500’N 30º20.333’W, 318 m deep, 30 August 1888).
Macroscopic description. Erect, flabellate sponges ( Fig. 6E–G View FIGURE 6 ; Table 5 View TABLE 5 ). They are 35–100 mm in height, with a basal stalk-like region in which the lamina progressively increases in wideness from the attachment point up to about 1/4 of the height, where it becomes approximately rectangular (3–9 mm in wideness). The lamina is thin (1– 1.5 mm) and flexuous. It can be undivided, or, in some individuals, forming two or three flattened branches, all with a regular apical margin ( Fig. 6E View FIGURE 6 ). The sponge surface is porous to the dissecting microscope, largely and irregularly hispid. Most collected individuals are pale orange when alive, turning into yellowish white to beige after ethanol preservation. Nevertheless, three of the specimens show remarkable color dissimilarity, being dark brown to black, at least after preservation in ethanol ( Fig. 6F–G View FIGURE 6 ). Some of these blackish specimens (part of the paratype series) also account for the largest sizes (up to 100 mm in height) and have the stalk-like region more flattened than the orange-beige individuals.
Skeleton. Megascleres are styles and oxeas ( Table 5 View TABLE 5 ). Styles occur in a wide variety of size and shape, with abundant intermediate forms that prevent making putative categories. Styles are often slightly curved, either symmetrically or asymmetrically; sometimes they have more than one flexion point and can even be angulated and, more rarely, slightly sinuous ( Fig. 8A–C View FIGURE 8 ). The round end is usually regular, occasionally with an annular swelling ( Fig. 8A View FIGURE 8 ). The distal end is of variable morphology, from sharp hastate or acerate type to stepped, mucronate, and almost blunt (strongyloxea-like) type ( Fig. 8A View FIGURE 8 ). Styles measure 119– 1400 x 3–30 µm, not being further categorizable according to their size. They are also difficult to separate according to their location, but most of those in the choanosomal region are not larger than 550–620 x 10–20 µm. Styles in the black specimens have a size range (245– 1400 x 5–20 µm) virtually identical to that of the orange-beige individuals, although predominating sizes are usually over 800 x 10 µm. Oxeas, some more abundant than the styles, are also relatively variable in size and shape, but variability ranges are similar among specimens. They can be slightly or markedly curved, once or twice, and symmetrically or asymmetrically ( Fig. 8A, D View FIGURE 8 ). Tips are usually acerate or blunt, although mucronate ends also occur ( Fig. 8A View FIGURE 8 ). They measure 180–750 x 2.5–20 µm in the orange-beige specimens and 120–500 x 2.5– 20 µm in blackish individuals. Microscleres in both orange-beige and blackish individuals are raphides in highly packed trichodragmata ( Fig. 8E–F View FIGURE 8 ), measuring 22.5–35 x 5–13.7 µm. Although no evident size difference exists in trichodragmata between orange-beige and blackish individuals, their shape can be flattened or cylindrical in the orange-beige ones ( Fig. 8E View FIGURE 8 ), but only cylindrical trichodragmata ( Fig. 8F View FIGURE 8 ), and in higher abundance, are found in the blackish ones.
The skeletal structure is plumoreticulate. There is an evident axial skeleton in the stalk-like region, built by multispiculate tracts of oxeas. In the thin lamina there is no clear distinction between axial and extra-axial skeleton. Rather, there are pauci—and multispicular, ascending and ramifying tracts, compressed in the sense of the lamina and consisting of mainly oxeas, with sparse styles ( Fig. 8G View FIGURE 8 ). These tracts are looser than in the axial skeleton of the stalk, and are connected each other by an irregular reticule that becomes more apparent in the thinnest parts of the lamina . In the three blackish specimens, the styles in the ascending tracts are somewhat more abundant and usually slightly longer than those in the orange-beige specimens. There are peripheral styles with their round end embedded in the tracts, piercing perpendicularly surface and making it hispid. The hispidating styles are usually long and occur isolated or in plumose tufts of up to 7 styles. Trichodragmata are predominantly located near the surface, especially in the blackish specimens. Spongin is abundant in the axial skeleton although it does not entirely embed all the spicules. It occurs moderately in the plumose tracts of the lamina ( Fig. 8G View FIGURE 8 ).
Distribution and ecology notes. All the collected specimens came from gravel bottoms, sometimes with organogenic components, at depths ranging from 93 to 173 m.
Taxonomic remarks. Except for color, the morphological and skeletal differences between the orange-beige individuals and the blackish ones are minor ( Table 5 View TABLE 5 ) and we judged them not enough to support a differentiation into two separated species. Both color varieties share relevant features, such as similar body morphology and plumoreticulate skeleton with the same spicule categories and similar size ranges. In addition to the obvious color differences, it can be noticed: 1) a higher abundance of cylindrical trichodragmata in the blackish individuals; 2) a slightly more organized reticule-like arrangement linking the plumose tracts in the orange-beige individuals; and 3) higher frequencies of short styles in the orange-beige specimens. Even though we are herein assuming that these differences correspond to ill-known aspects of intraspecific variation, we cannot discard that future studies based on molecular features of new collections and/or "in vivo" observations may led to a species split.
Trichodragmata similar to those in A. spatula nov. sp. are also found in some other Axinella spp. (Table 3), such as Axinella infundibuliformis (Linnaeus, 1759) . Nevertheless, this latter species has a caliculate or fan-like body shape, a plumoreticulate skeleton of oxeas and styles clearly smaller (300–600 x 12–16 µm), as well as smaller (15–20 µm long) trichodragmata ( Lendenfeld 1897; Arndt et al. 1935). It is also worth noting that the earliest descriptions of A. infundibuliformis were little accurate and apparently overlooked the small trichodragmata ( Johnston 1842; Bowerbank 1866; Hansen et al. 1885; Fristedt 1887). Axinella alba ( Descatoire, 1966) also shows trichodragmata, but it is an encrusting species and has oxeas (700– 1000 x 15–18 µm) longer than those of A. spatula nov. sp. Trichodramata and styles also occur in Axinella flustra ( Descatoire 1966) , but, again, although these styles and trichodragmata are in a size range similar to those of A. spatula nov. sp., the branching body shape ( Fig. 6H–J View FIGURE 6 ) and shorter oxeas make A. flustra easily distinguishable (Table 3).
Some members of the axinellid genus Dragmacidon Hallman, 1917 (Table 3) bear some vague resemblance to A. spatula nov. sp., namely occurrence of raphides and the absence of a clear axial and extra-axial skeleton differentiation. Furthermore, phylogenetic analyses based on 18SrRNA, 28S rRNA and CO1 have brought Dragamacidon species and raphide-bearing Axinella species into a same clade ( Gazave et al. 2010; Morrow et al. 2012). Dragmacidon tuberosum ( Topsent, 1928) is the only geographically close species in the genus having trichodragmata, but those are distinctive, having the raphides projecting from each side of the packets; besides, this species has shorter styles ( Topsent 1928).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |