
Colossendeis sinuosa Stock, 1997
|
publication ID |
https://doi.org/ 10.5281/zenodo.157770 |
|
DOI |
https://doi.org/10.5281/zenodo.5694686 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87F7-6C65-553E-1B2A-F90CFE467C4A |
|
treatment provided by |
Plazi |
|
scientific name |
Colossendeis sinuosa Stock, 1997 |
| status |
|
Colossendeis sinuosa Stock, 1997 View in CoL
Stock, 1997, 398–399, fig. 4.
Material: New Caledonia: 1 specimen (MNHNPy 1005), station CP03, N/ O 'Alis' Campagne LITHIST Banc Stylaster, 23º27.0'S 167º41.5'E, 447 m; 10/08/1999, coll. Richer IRD. 1 specimen (NHM.2003.637), station CP1664, Ride de Norfolk. N/ O 'Alis' Campagne NORFOLK 1. Banc Stylaster, 23º40'S 167º43'E, 456–478 m; 20/06/2001, coll. Lozouet, Boissellier, RicherIRD. 1 specimen (MNHNPy 1018), station DW1694, Ride de Norfolk. N/ O 'Alis' Campagne NORFOLK 1. Banc Introuvable, 24º40'S 168º39'E, 575–589 m; 24/06/2001, coll. Lozouet, Boissellier, RicherIRD.
Solomon Islands: 1 specimen (MNHNPy 1033), station DW1741, N/ O 'Alis' Campagne SALOMON 1, 11º29.1'S 159º57.4'E, 557–655 m; 23/09/2001, coll. Bouchet, Dayat, Warén, RicherIRD.
Additional specimen examined: 1 specimen (MNHNPy 881), station CP32, 23º38'S 167º43'E, 420–460 m; 18/10/1992, identified as C. pipetta by Stock (1997).
Remarks: Stock (1997) distinguished this species from C. pipetta on the basis of its pipettelike proboscis tip being sinuous rather than regularly downturned, in its palp articles being shorter, and in its tarsus being only 1.5 times the length of the propodus. The species was described from 4 specimens (three from a single sample) from New Caledonia at 560– 730 m. The present material represents only the second time the species has been recorded.
With a number of specimens from a range of sites and depths, it was considered appropriate to undertake some more detailed analysis of the morphometrics by which Stock (loc. cit.) had distinguished the two species. Analyses were based on the present material, the detailed measurements of the types of both species, plus examination of a number of paratypes kindly loaned by the Muséum National d'Histoire Naturelle, Paris (15 specimens, registration numbers MNHNPy881, 884, 886, 887, 889–891, 893 and 894).
Measurements were made on each of a total of 35 specimens (including four putative C. sinuosa ) using a curtain micrometer eyepiece, with accuracy to 0.005 mm. Measurements were made of the trunk length (from the anterior edge of the cephalon to the tip of the fourth lateral process), the lengths of the tarsus and propodus of the third leg (where possible) and of palp articles 4 to 7.
No significant distinctions were found from the proportions of palp articles, their length ratios were consistent over the size range (3.75 to 8.59 mm trunk length) of the animals studied, as was their ratio to the trunk length itself. Thus, Stock’s contention of a distinction of these species on the basis of palp article proportions is not supported.
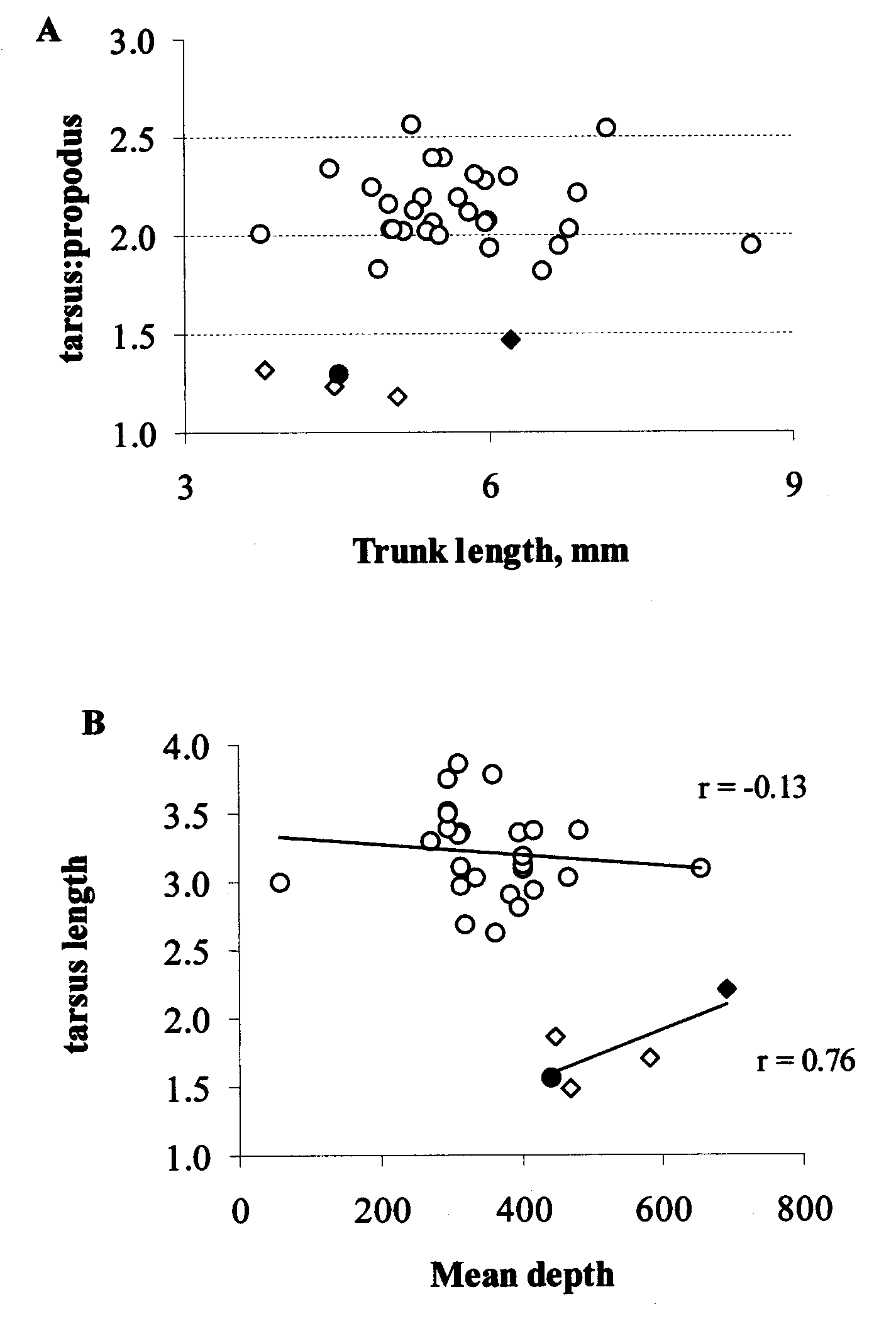
The tarsus showed no significant proportionate trend with size of the animal, while propodus became proportionately significantly shorter in larger animals (r = 0.86; p <0.001). The ratio of the tarsus to the propodus did split the data into two groups ( Fig. 7 View FIGURE 7 A); the ratio showed no correlation with propodus length but increased in significant positive relationship to tarsus length (r = 0.91; p <0.001). Tarsus length on its own consistently distinguished five specimens, including the holotype of C. sinuosa , those specimens listed above attributed to C. sinuosa from the New Caledonia samples, and one of Stock’s (1997) C. pipetta specimens (MNHNPy 881). These specimens had a tarsus to propodus ratio of 1.47 or less, whereas all the other (putative C. pipetta ) material had a ratio greater than 1.8 (1.82 to 2.56). As stated above, the distinction by this ratio is caused by the differential length of the tarsus, not the propodus.
There was some indication of a decline in this ratio with depth (or a deeper distribution of C. sinuosa ). However, when the relationship is tested for each species separately, it is seen that their trend with depth is neither significant nor consistent between the two ( Fig. 7 View FIGURE 7 B).
It is concluded that the ratio of the tarsus to the propodus is a valid parameter by which to distinguish these two species, with Colossendeis sinuosa having a tarsus to propodus ratio of 1.5 or less, C. pipetta a ratio of 1.8 or more (and thus specimen MNHNPy881 is of the former species, collected from very close to the site of one present specimen from station CP1664).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |