
Pseudotomentella sorjusensis Svantesson, 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.497.2.1 |
|
DOI |
https://doi.org/10.5281/zenodo.6495788 |
|
persistent identifier |
https://treatment.plazi.org/id/03D787D3-1633-F968-FF4A-14B3D26308F8 |
|
treatment provided by |
Marcus |
|
scientific name |
Pseudotomentella sorjusensis Svantesson |
| status |
sp. nov. |
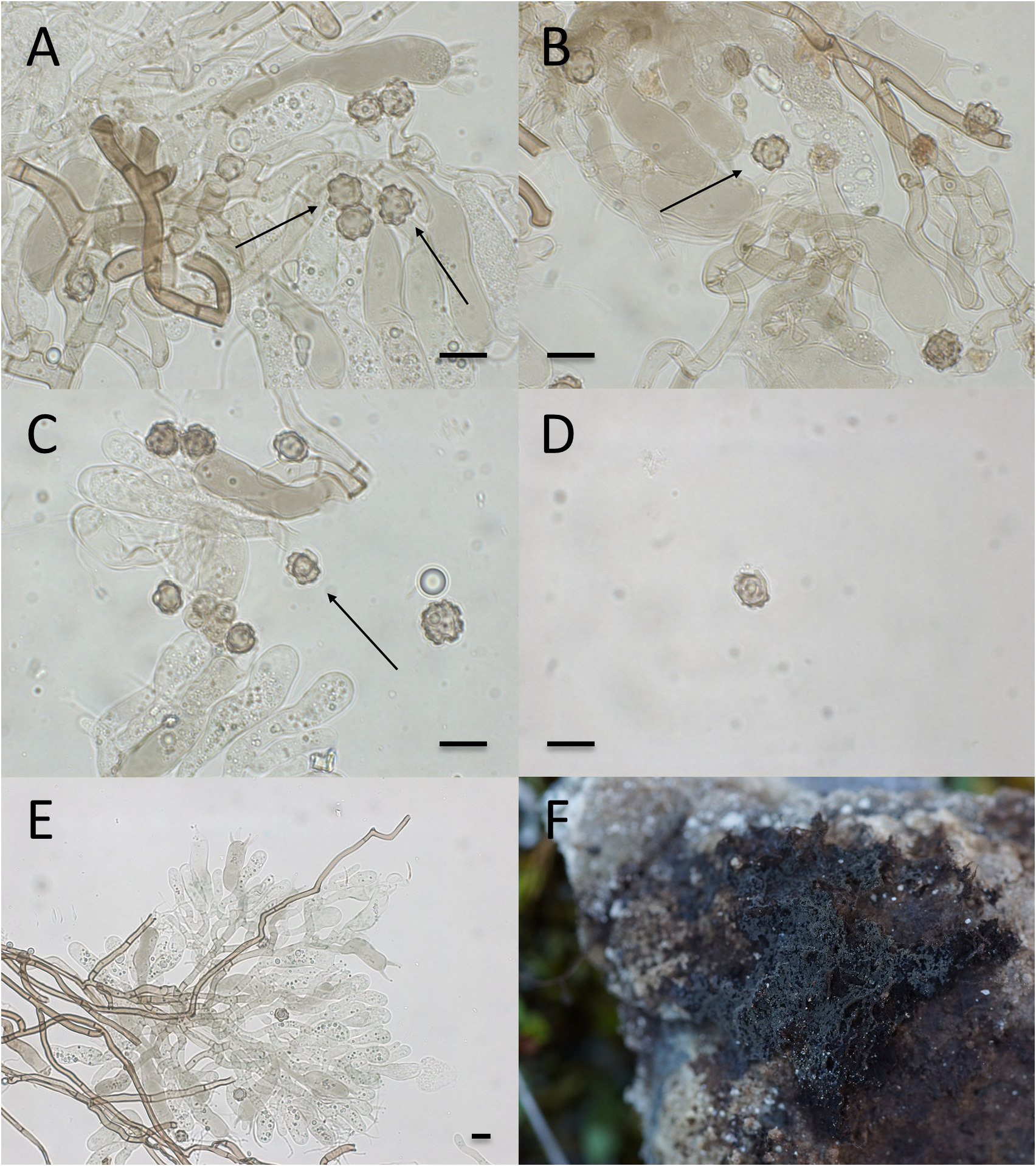
Pseudotomentella sorjusensis Svantesson View in CoL , sp. nov. ( Fig. 4 View FIGURE 4 )
MycoBank No.: MB 835163 . UNITE SH: SH1185284.08 FU. Etymology: the name refers to Sorjus, an older spelling of the type locality .
Type: SWEDEN. Lule Lappmark : Jokkmokk, Sårjås N , low alpine heath on ground with intermediate pH, on underside of stone, 17 August 2016, S . Svantesson 298 (holotype: GB!, GenBank Acc. No. ITS: MT 146448 View Materials ) .
Basidiome annual, resupinate, membranaceous; effused to approximately five centimetres in diameter. Mature parts continuous, with a firm, fibrous and compact yet soft and rather elastic texture. Hymenium smooth; greenish brown when fresh, brown with a reddish hue when dried. Immature parts discontinuous, byssoid with a cottony texture. Subhymenium and hymenium of immature parts blue grey when fresh, blue grey to grey brown when dried. Subiculum well-developed, loose, fibrous, brown; forms the outer edge of the basidiome, extending noticeably beyond the hymenium.
Hyphal cords lacking, but loose bundles of subicular hyphae sometimes present.
Hyphal system monomitic, clamp connections absent from all hyphae.
Subicular hyphae noticeably long and straight, thick-walled; forming a loose tissue. Individual hyphae (3.0–) 3.1–4.3 μm wide, with a mean width of 3.6 μm; brown to orange brown in KOH, orange brown in water; inamyloid.
Subhymenial hyphae often somewhat sinuous, thin to thick-walled; forming a rather dense tissue. Individual hyphae 3.3–5.2 (–5.5) μm wide, with a mean width of 4.3 μm; hyaline to brown in KOH, with a green or blue green reaction in the presence of air; pale green to pale orange green in water, with strongly granular contents; occasionally amyloid.
Encrustation none observed.
Basidia with four slightly curved sterigmata, occasionally two-sterigmate; clavate to narrowly clavate, sometimes clavopedunculate, thin-walled, with one–three slight constrictions. Dimensions: 41–56 (–59) × (10.1–) 10.3–12.1 (– 12.6) μm; mean dimensions: 48 × 11.5 μm. Sterigmata (7.0–) 7.2–8.9 (–9.2) μm long, with a mean length of 8.0 μm. Colours and reactions the same as for subhymenial hyphae; amyloid reaction most frequently found at the bases of basidia.
Cystidial organs lacking.
Basidiospores in frontal face generally with a subcircular or triangular basic shape and an angular, nodulose, triangular or sometimes cross-shaped outline, covered in bi-or trifurcate, sometimes singularly attached, echinuli. A majority of the spores with three-five distinct, rounded to square lobes; seven-lobed spores occasionally occurring; abnormally large spores originating from two-sterigmate basidia infrequently seen. Frontal dimensions: 7.4 – 8.6 (–9.1) × 7.7–8.8 (–9.1) μm; mean dimensions: 8.1 × 8.2 μm; Q-value: 0.9–1.1; mean Q-value: 1.0. Echinuli (0.5–) 0.6–0.8 μm long, with a mean length of 0.7 μm. Lateral face ellipsoid to ovoid, with evenly rounded edges or one–three lobes. Lateral dimensions: 7.4–8.5 × (5.0–) 5.2–6.3 (–6.5) μm; mean dimensions: 7.9 × 5.8 μm; Q-value: 1.2–1.5; mean Qvalue: 1.4. Colour in KOH pale brown to pale orange brown, in the presence of air sometimes with a green to blue green reaction; in water pale orange brown; occasionally amyloid.
Chlamydospores lacking.
Habitat
The only specimen recorded to date of P. sorjusensis is the type collection, which was found in a low alpine heath on ground with intermediate pH. UNITE sequence metadata show that the species forms ectomycorrhiza with at least Picea abies (L.) H. Karst., Picea glauca (Moench) Voss , Salix arctica Pall. and Salix caprea L. ( Kõljalg et al. 2005, Nilsson et al. 2018). One of the root tip sequences originate from an arctic locality, while the remaining sequences in the UNITE SH come from temperate forests in lowland areas.
Distribution
Basidiomata encountered in: Sweden. Root tip samples confirm presence also in Estonia (3), Canada (2), and soil samples in Estonia (56) and Latvia (2).
Remarks
Within the P. tristis group, the basidiome of P. sorjusensis can be recognised by its lack of hyphal cords and skeletal hyphae, its dense, compact texture after drying, bluish colour of immature parts, narrow subicular hyphae and its short spores. Two species, P. badjelanndana and P. rotundispora are similar to P. sorjusensis . Pseudotomentella badjelanndana has thinner subhymenial hyphae, whose mean diameter is smaller than its subicular hyphae. Its spores are also generally longer than wide and have longer echinuli but a larger frontal face than in P. sorjusensis . Pseudotomentella rotundispora differs from P. sorjusensis by slightly thinner subhymenial hyphae, which are of more or less equal width to its subicular hyphae and by its spores, which are slightly shorter in frontal face. For further notes on the morphological separation of species within the P. rotundispora group see Remarks under the description of P. badjelanndana . Other described species within the group can appear similar, but have either wider hyphae, longer spores or both.
| MB |
Universidade de Lisboa, Museu Bocage |
| SH |
Academia Sinica |
| FU |
Fudan University, Department of Biology |
| N |
Nanjing University |
| S |
Department of Botany, Swedish Museum of Natural History |
| GB |
University of Gothenburg |
| MT |
Mus. Tinro, Vladyvostok |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |