
Cladorhiza investigator, Ekins & Erpenbeck & Hooper, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4774.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B0C4A2F8-F2AB-4147-BB12-63720EEF2516 |
|
DOI |
https://doi.org/10.5281/zenodo.3846437 |
|
persistent identifier |
https://treatment.plazi.org/id/3D3FEFCD-742A-42A2-BCD4-E7B559608EE5 |
|
taxon LSID |
lsid:zoobank.org:act:3D3FEFCD-742A-42A2-BCD4-E7B559608EE5 |
|
treatment provided by |
Plazi |
|
scientific name |
Cladorhiza investigator |
| status |
sp. nov. |
Cladorhiza investigator View in CoL sp. nov.
Figures 12 View FIGURE 12 & 13 View FIGURE 13 , Table 5 View TABLE 5
urn:lsid:zoobank.org:act:3D3FEFCD-742A-42A2-BCD4-E7B559608EE5
Material examined: Holotype: QM G337458 off Freycinet Peninsula , Station 11, Tasmania, Australia, 41° 43’ 14.5” S, 149° 7’ 30.7” E, 2793 m, Box Corer, Coll. Merrick Ekins on RV Investigator, Cruise IN2017_ V03 , Sample 11-103, 19/v/2017. GoogleMaps
Etymology: Named for the Marine National Facility’s research vessel Investigator, as a noun in apposition, used to sample the bathyal and abyssal fauna of Eastern Australia.
Distribution: This species is only known so far from the bathyal type locality off the Freycinet Peninsula, Tasmania, Australia, at bathyal depth.
Description:
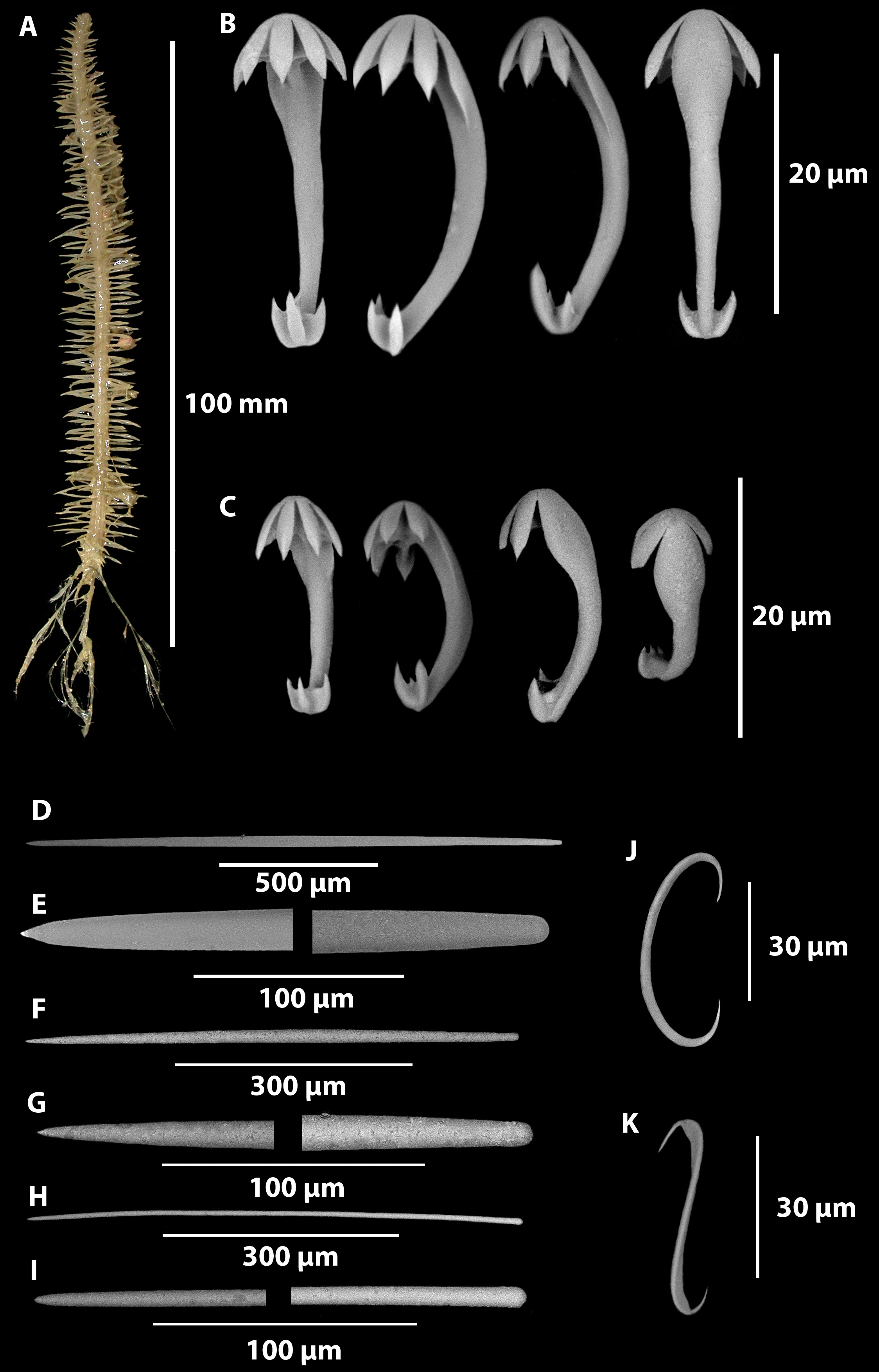
Growth form: A stipitate erect, unbranched sponge with horizontal filaments at right angles and encircling the entire stem (body) ( Figure 12 A View FIGURE 12 ). These lateral filaments form up to nine distinct columns, the largest are up to 8 mm long at the base of the stem, the smallest 2–3 mm long at the apex of the stem, 0.18–0.2 mm wide, and spaced 1.7–2.5 mm apart.. The sponge stem (body) is 92 mm long and 2.9 mm wide, with basal rootlets, the longest being approximately 25 mm long, branching one or more times along their length. Total body length including rootlets and stem is 117 mm long
Colour: Cream on deck and in ethanol.
Ectosomal skeleton: The ectosomal skeleton of the filaments is thin (~ 35 µm), membranous, and contains anisochelae and sigmas ( Figure 13 A View FIGURE 13 ). The ectosomal skeleton of the stem is a thin membranous covering of a palisade of mycalostyles form very effective wall plates. ( Figure 13 C View FIGURE 13 ). The ectosomal skeleton of the roots is just a very thin membrane.
Endosomal skeleton: The axis of the stem has a radial skeleton, with a small core of confused small mycalostyles. From this nucleus nine arms radiate outwards ( Figure 13 E View FIGURE 13 ), composed of mycalostyles, forming the filaments. In between these radiating arms there are also seven radiating wedges of longitudinal mycalostyles supporting the stem of the sponge ( Figure 13 D View FIGURE 13 ). The outer edge of the wedge forms the Palisade-like wall of the ectosome.
Between these longitudinal wedges and the filament arms are thin wedges of tissue containing anisochelae and sigmas. The endosome of the filaments consists of tightly bound mycalostyles longitudinally arranged ( Figure 13 B View FIGURE 13 ). The endosome of the roots of the sponge, are composed of tightly bound bundles of mycalostyles and rare subtylostyles ( Figure 13 F View FIGURE 13 ).
Megascleres: Mycalostyles can be split into two size classes: large mycalostyles (1240-(1509)- 1880 x 18.3- (26.6)-40.9 μm, n=80) ( Figure 12 View FIGURE 12 D-E) and medium mycalostyles (600-(914)- 1210 x 11.6-(15.6)-21.2 μm, n=63) ( Figure 12 View FIGURE 12 F-G). Both styles occur in the filaments, stem and roots of the sponge. However the smaller of the mycalostyles also have an important supporting function in the buttressing of the filaments as they emerge from the stem ( Figure 13 D View FIGURE 13 ). In the roots and lower stem there are also thin subtylostyles (356-(624)-905 x 2.7-(6.2)- 11.6 µm, n=37) ( Figure 12 View FIGURE 12 H–I).
Microscleres: Multidentate unguiferate anchorate anisochelae, with five alae on the head and a much smaller foot bearing three alae, occurring in two significant size classes, with the larger category showing slightly different shape of the upper alae compared to the lower alae: large (20.3-(24.4)-27.8 x 1.1-(1.8)-3.5 μm, n=175), and small (13.2-(16.6)-19.1 x 0.8-(1.3)-2.7 μm, n=86) ( Figure 12 View FIGURE 12 B-C). There are uncommon sigmas which display none to only slight contortion (36-(79)-152 x 1.7-(3.4)-7.1 μm, n= 49) ( Figure 12 J View FIGURE 12 ). There are also rare sigmancistras showing 90 o contortion (36-(40)-44 x 1.6-(2.1)-2.7 μm, n=9) ( Figure 12 K View FIGURE 12 ). Both the sigmas and sigmancistras are located in the ectosome of the filaments and the endosomal wedges of the stem.
Molecular data: The 28S sequence of QM G337458 is provided in the Sponge Barcoding Database under accession number SBD#2310 and the molecular difference to other congenerics displayed in Figure 3 View FIGURE 3 .
Remarks: Cladorhiza investigator sp. nov. is most importantly different to other closely allied species by the presence of tylostyles in the roots and base of the stem. Cladorhiza investigator sp. nov. has a similar growth form, spicule types, and approximately the same spicule sizes as the holotype of C. rectangularis ( Ridley and Dendy, 1887) , from the central South Pacific. Those authors only mention one of each type of spicule and give only one measurement as was usual at that time of publishing, with one type of style, one type of anisochelae and one type of sigma mentioned, and indeed they also mention the fragility of the dried holotype. However, Koltun (1959, 1970) described additional specimens which he assigned to C. rectangularis from the North and Northwest Pacific, respectively, both with similar spicule diversity and measurements to those given by Ridley & Dendy (1887) ( Table 5 View TABLE 5 ). Cladorhiza investigator sp. nov. shows some differences in morphology to C. rectangularis most notably the number of rows of filaments encircling the stem (nine rows in the new species, compared to 4 rows in 2 vertical planes in C. rectangularis ), and the length of filaments (8 mm compared to 17 mm, respectively), with the new species also having a thicker stem, and a tapered (non-rectangular) shape. Cladorhiza investigator sp. nov. also has two size classes of multidentate unguiferate anisochelae (the larger anisochelae also show slightly different dentition between the upper and lower alae), and only one size class each of sigmas and sigmancistras, whereas C. rectangularis has only one size of tridentate anisochelae, two sizes of sigmas, and lacking sigmancistras. The dimensions of the anisochelae and sigmas also differ between the two species ( Table 5 View TABLE 5 ).
Cladorhiza evae ( Lundsten et al., 2014) from the bathyal off Cabo Pulmo, Mexico, also closely matches the description of C. investigator sp. nov. It has a gross morphology more similar to that of C. rectangularis (rectangular non-tapering body and filaments), than to the new species, with four to five columns of filaments (compared to four in C. rectangularis and nine in the present species). Lundsten et al. (2014) argue that the differences between C. evae and C. rectangularis include greater style width, larger sigmas of two sizes classes, and the presence of sigmancistras in the former. Also similar to the new species, C. evae has mycalostyles split into three size classes, with the smaller category more abundant in the filaments but also occurring in the axial skeletons of the stem and basal rhizoid rootlets.
Other differences between the C. investigator sp. nov. and C. evae include the possession of two categories of unguiferate multidentate anisochelae and only one category of sigmas in the former, compared with one and two categories, respectively, in the latter species.
Nevertheless, C. investigator sp. nov., C. evae and C. rectangularis are clearly sister morphospecies, and their molecular taxonomic differences are also reflected by molecular data (see on the sequence of the paratype of C. evae CASIZ192775, Fig. 3 View FIGURE 3 ). Population genetic data is required to explain the pan-Pacific distribution of this species group.
Göcke & Janussen (2013) provide a good discussion of similar species including Cladorhiza penniformis Göcke & Janussen, 2013 , but that Antarctic species is described as feather-like with the filaments predominantly in one plane, and a slightly thinner and flattened stem. This new sponge species is also very similar to C. caillieti ( Lundsten et al., 2014) , however, again, it has almost twice as many rows of lateral processes and has different shaped anisochelae.
Cladorhiza segonzaci Vacelet, 2006 also has some similarities to the new species; however, the new species has a second larger size category of anisochelae, and larger styles than C. segonzaci . It also vaguely resembles Lycopodina occidentalis ( Lambe, 1893) , but of course has very different microscleres.


| QM |
Queensland Museum |
| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |