
Spio filicornis ( O . F. Müller, 1776 ), O. F. Muller, 1776
|
publication ID |
https://doi.org/ 10.5281/zenodo.277166 |
|
DOI |
https://doi.org/10.5281/zenodo.5697053 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF3732-FFC4-FFD4-39E8-FE8B8384317C |
|
treatment provided by |
Plazi |
|
scientific name |
Spio filicornis ( O . F. Müller, 1776 ) |
| status |
|
Spio filicornis ( O. F. Müller, 1776) View in CoL
( Figs 1 View FIGURE 1 , 3, 4, 5, 6, 7A, C, 8A)
Nereis filicornis O.F. Müller, 1776: 218 . – Fabricius, 1780: 307 –308
Spio filicornis ( O.F. Müller, 1776) View in CoL . – Fabricius, 1785: 264 –269, plate V, figs. 8–12.
Neotype. West Greenland, Iluilârssuk, near Paamiut (Frederikshaab): 61° 50.059N, 49° 25.183W, 14.06.2009, eulittoral (ZSRO P-2059).
Non– type material. West Greenland, Iluilârssuk, near Paamiut (Frederikshaab): 61° 50.059N, 49° 25.183W, 14.06.2009, eulittoral: 16 specimens ( ZSRO P- 2048), 1 specimen ( ZSRO P- 2056), 1 specimen ( ZSRO P- 2057), 1 specimen ( ZSRO P- 2058), 5 specimens ( ZSRO P- 2060), 1 specimen ( ZMH P- 25869), 3 specimens ( ZMH P- 25870), 1 specimen ( SMF 19520), 3 specimens ( SMF 19521); West Greenland, Paamiut: 62° 00.709N, 49° 41.208W, 12.06.2009, eulittoral: 1 specimen ( ZSRO P- 2009), 11.06.2009, 3 specimens ( ZSRO P- 2044).
Diagnosis. Anterior part of prostomium bluntly rounded, posterior part with high, narrow, keel-shaped elevation; anterior part of prostomium and peristomium with dark brown pigmentation; palps with dark brown spots along posterior margin of food groove; nuchal organs with two pairs of more or less straight ciliated bands posterior and posterolateral to prostomium before 2nd transverse ciliated bands (tcb) on chaetiger 3; branchiae of chaetiger 1 shorter and narrower than those on following chaetigers, absent on last six to ten chaetigers; double-paired metameric dorsal ciliated organs present between tcb of two consecutive segments, starting between tcb of chaetigers 3 and 4, i.e., after second tcb, continuing up to tcb between chaetigers 10 and 11, i.e., up to tenth tcb (rarely up to 11th tcb on chaetiger 12); neuropodial postchaetal lamellae on median chaetigers narrow but distinctly elongated dorsally; notopodial postchaetal lamellae on posteriormost abranchiate chaetigers elongated lobe-like, distinctly longer than wide.
Description. Neotype complete specimen (without palps) with 52 chaetigers, about 15 mm in length and 1.4 mm wide. Other examined specimens with 41–60 chaetigers, 9–20 mm in length and 1–2 mm in width.
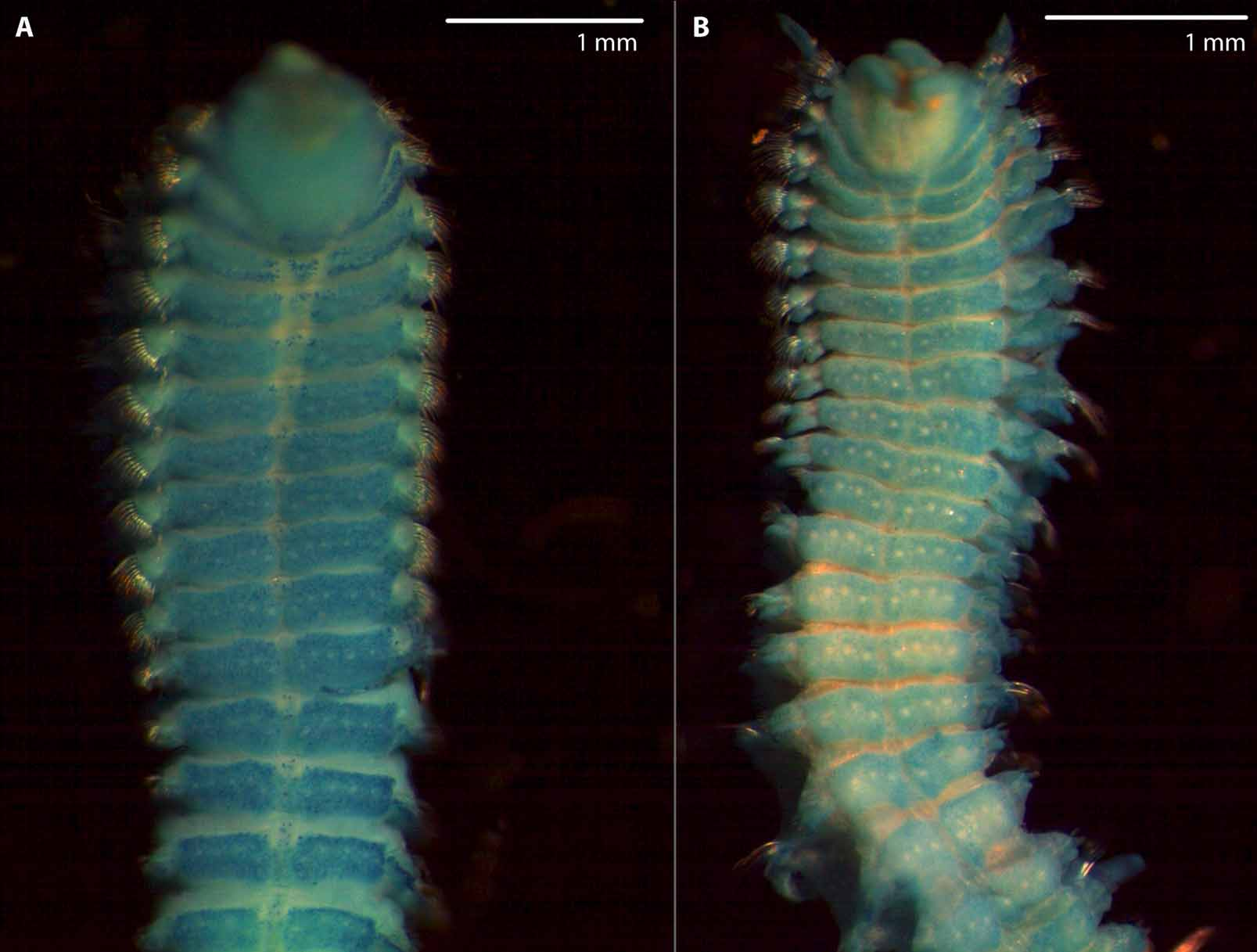
Anterior part of prostomium anteriorly rounded, sometimes with slight median incision (Figs 3A, 5A, C, 7A); posterior part of prostomium with high, narrow, keel-shaped elevation, beginning in front of anterior pair of eyes and terminating on chaetiger 1 (Figs 3B, 5C, 6C, 7C); transverse depression between anterior and posterior part of prostomium (Fig. 3B); 2 pairs of black eyes arranged in trapezoid, anterior pair crescent-shaped or oval, widely spaced, posterior pair oval or round, closely spaced (dark brown spots of pigment between anterior and posterior eyes easily mistaken as a third pair of eyes) (Fig. 3A); prostomium distinctly separated from peristomium by furrow; peristomial palps comparatively short, reaching chaetiger 6 to 8 at maximum ( Fig. 6 View FIGURE 6 ).
Nuchal organs with two pairs of more or less straight ciliated bands up to second tcb on chaetiger 3; median pair of ciliated bands interrupted by 1st tcb on chaetiger 2: anterior parts of median ciliated bands from just posterior to prostomium to 1st tcb, posterior parts of median ciliated bands posterior to 1st tcb; pair of lateral ciliated band outwards curved in the range of first tcb; median and lateral ciliated bands not connected in front of second tcb (Figs 3A, 5A, C); metameric dorsal ciliated organs as double-paired ciliated bands after 2nd tcb, usually up to 10th tcb on chaetiger 11 (rarely up to 11th tcb on chaetiger 12) (Figs 3A, 5A, D, 6B, 7A).
Branchiae from chaetiger 1, continuing to almost the end of body, only last six to ten chaetigers without branchiae (neotype: seven chaetigers without branchiae); branchiae on first chaetiger about one-third shorter and narrower than those on following chaetigers (Figs 3A, B, 5C), reaching midline dorsally and touching on some medium chaetigers, becoming thinner and shorter posteriorly: last two to five branchiae distinctly shorter than notopodial postchaetal lamellae (Fig. 5E); branchiae with narrow base, tapering distally, with cilia on inner and a furrow on outer side (Figs 5B, D); branchiae on anterior chaetigers basally fused with notopodial postchaetal lamellae, separated from lamellae in posterior chaetigers (Fig. 4).
First notopodium shifted dorsally. Notopodial postchaetal lamellae on anterior chaetigers oval, base of postchaetal lamellae longer than transverse row of corresponding chaetae in anteriormost chaetigers; notopodial postchaetal lamellae becoming smaller and rounded on medium chaetigers, elongated lobe-like on posterior chaetigers, straighten up on very last chaetigers without branchiae (Figs 3B–E, 4, 5B, E). Neuropodial postchaetal lamellae in anterior chaetigers rounded, becoming smaller, elongated and notched from about chaetiger 12 to 14, rounded in posterior chaetigers (Figs 3B–D, 4, 5B). Notopodial and neuropodial postchaetal lamellae may touch each other (on chaetigers 17–19 in neotype) or even interconnected on some anterior or mid-body chaetigers (Figs 4C, 5B).
Notopodial chaetae all capillaries with narrow sheath; capillaries of anterior chaetigers arranged in two rows: chaetae of anterior row short, broad, distinctly granulated; chaetae of posterior row longer, thinner, lacking granulations; additional superior fascicle of very long, thin capillaries, without granulations; capillaries of posterior chaetigers arranged in irregular rows, thin, non-granulated, of different lengths. Neuropodia with rows of capillaries and hooded hooks as well as an inferior fascicle of capillaries; capillaries of anterior neuropodia arranged in two rows, similar to notochaetae (Fig. 3I); posterior row replaced by single row of 5 to 9 (5 to 6 in neotype) hooded hooks from chaetiger 11 (Fig. 3H); on posteriormost chaetigers a third row of very thin alimbate fine-granulated capillaries (Fig. 3F); hooks not narrowed subdistally, with a comparatively long hood, bidentate, main fang and apical tooth well developed; inferior fascicle with 2 to 5 long, thin, limbate capillaries without granulations from chaetiger 1, replaced by 1 to 2 sabre chaetae from medium chaetigers, each distally granulated (Fig. 3G).
Pygidium with four anal cirri of about equal length and width; dorsal pair more widely spaced than ventral pair. Anus usually dorsally shifted (Figs 3D, E).
Pigmentation. Palps on live specimens with reflecting whitish and light brown spots, giving palps a ringed appearance ( Fig. 6 View FIGURE 6 A); pigmentation fades in ethanol and formalin but light brown pigment along food groove remains, posterior margin of food groove additionally with about 8–15 dark brown spots along total length of palps ( Figs 6 View FIGURE 6 B, C); alcohol-preserved specimens with light and dark brown pigment on anterior part of body as follows: prostomium anterior to transverse depression with light to dark brown pigment; peristomium heavily pigmented, especially on median (adjacent to prostomium) and posterior margin (Figs 3A, 7A); pigmented patches in front of and in particular behind tcb dorsolaterally from chaetiger 2 to about chaetiger 10 or 12 (chaetigers 2–10 in neotype) (Figs 3A, 7A); dark brown pigment between and around noto- and neuropodial lamellae, particularly distinct on first two or three chaetigers (distinct pigmentation present on chaetigers 1–10 in neotype) (Figs 3B, 6A, 7C); anterior and posterior margin of branchial base from chaetiger 1 to about 10 with dark brown pigment (chaetigers 1–9 in neotype) ( Fig. 7 View FIGURE 7 A); sometimes keel-shaped elevation of prostomium also with light brown pigment; dark brown spots of pigment between anterior and posterior eyes, sometimes light brown pigment close to eyes; no pigment ventrally apart from anterior margin of prostomium and median margins of peristomium.
Methyl green staining pattern. Anterior part of prostomium and caruncle intensively stained; epidermis middorsally in front of and in particular behind tcb from about chaetiger 5 to chaetiger 25, and laterally between neuro- and notopodium from chaetiger 7 to chaetiger 18 intensively stained; margins of postchaetal lamellae and branchiae of anterior and medium chaetigers intensively stained; in posterior chaetigers postchaetal lamellae and branchiae completely deep blue after staining.
If specimens transferred from the methyl green solution into distilled water again, after about 10 minutes 1 to 6 pairs of white dots against the bluish background become visible ventrally on anterior chaetigers ( Fig. 8 View FIGURE 8 A). First pair of dots usually visible on chaetiger 4, 5 (rarely 6) pairs are present from chaetiger 8 to chaetiger 12, diminishing continuously to one pair on chaetigers 20 to 25, rarely chaetiger 27 (up to chaetiger 23 in neotype). The following pattern of white dots were found on chaetigers 4–20 on the investigated specimens (n=18, data for the neotype in parentheses): chaetiger 4: 1 (1), 5: 2 (2), 6: 3 (4), 7: 4 (4), 8–12: 5 (5), 13: 4 (5), 14–15: 4 (4), 16: 3 (4), 17–19: 2 (3), 20: 1 (3). These dots are arranged in a row but the median pair is shifted slightly posteriorly. They obviously correspond to the ventral openings shown in Fig. 5F (see discussion).
Biology. Spio filicornis was found in intertidal and shallow subtidal regions of sandy beaches and in sandy sediments of rockpools between stones, gravel and macrophytes ( Fucus sp.). Recently settled juveniles with about 20 to 30 chaetigers have also been found in the habitat of the adults in June. The habitat of S. filicornis is strongly influenced by ice in winter and melt water in summer. A lot of peritrichous ciliates were attached to the anterior part of the trunk especially on the dorsal side adjacent to the tcb and on the branchiae (Figs 5C, C’). These epibionts were also found on juveniles.
Geographical distribution. So far known only from Iluilârssuk (type locality) and Paamiut (Frederikshaab), West Greenland.
FIGURE 3. Spio filicornis ( O.F. Müller, 1776) : A. Anterior end, dorsal view. B. Anterior end, lateral view. C. Chaetigers 9–13, lateral view. D. Posterior end, lateral view. E. Posterior end, dorsal view. F. Posterior neurochaeta from parapodium of the 9th last chaetiger. G. Sabre chaeta from 18th last chaetiger. H. Neuropodial hooded hook from the 9th last chaetiger. I. Anterior neurochaeta from the same.— All from type locality (ZSRO-P2060); Scale: A–C 0.5 mm, D 0.4 mm, E 0.3 mm, F, G, I 5 µm, H 10 µm.
FIGURE 4. Spio filicornis ( O.F. Müller, 1776) : A–F. Parapodium from chaetiger 1, 10, 11, 20, 9th last, and 18th last, all anterior view. — All from type locality (ZSRO-P2060); Scale: A–F 0.1 mm.
FIGURE 5. Spio filicornis ( O.F. Müller, 1776) : A. Anterior end, dorsal view; palps lost. B. Chaetiger 10–16, lateral view; arrow points at dorsally extended neuropodial lobe in 13th parapodium. C. Anterior end, dorsal view, palps lost; upper white arrow points at keel-shaped elevation on the prostomium, lower white arrow points at one of the numerous epibionts commonly found on the cuticula of S. filicornis (see C´); lcb=(left) lateral ciliated band of the nuchal organ, mcb=(left) median ciliated band of the nuchal organ. C’. Epibiont (Ciliata) enlarged. D. Dorsal transverse ciliated bands (=tcb) of chaetigers 9–12 with dorsal ciliated organs visible on dorsum; arrows point at metameric dorsal ciliated organs. E. Posterior end, dorsal oblique view, left side destroyed; arrow indicates last papilliform branchiae. F. Ventrum of chaetigers 8–11 with segmental pattern of tiny holes corresponding with white dots in methyl green staining pattern, arrow points at one of the holes— A–D from type locality (ZSRO-P2060), E specimen from more northern location: Ilulissat, F specimen from Spitzbergen.
Remarks. Spio filicornis can no longer be regarded as a species of world-wide distribution. The distribution might be wider than mentioned above but this still needs to be investigated. Characters formerly used to identify S. filicornis have turned out to be inappropriate. Among the characters described above, the beginning and the posterior extension of the dorsal ciliated organs, the number of posterior chaetigers with short and without branchiae, the shape of the neuropodial lamellae on midbody chaetigers and the appearance and number of white dots have been found to be significant for this species in particular (see also remarks on S. arndti sp. nov.). These dots are very likely the same structures referred to as ventral subepidermal glands by Maciolek (1990) in her description of S. filicornis from subtidal regions of the western North Atlantic (Maciolek, personal communication). However, those specimens examined by Maciolek (1990) obviously do not belong to S. filicornis since they exhibit a different pattern of white dots (6 dots per segment opposed to 10 or rarely 12), specimens are more slender (1 mm vs. 2 mm) and the first pair of branchiae is longer in comparison to S. filicornis from Greenland. Moreover, the nuchal organ is of a different shape according to the illustrations provided (see Maciolek 1990, Fig. 1 View FIGURE 1 B). No information on the dorsal ciliated organs is given.
Three species, Spio readi Blake, 1984 , S. malmgreni Sikorski, 2001 and Spio arndti sp. nov. are morphologically similar to S. filicornis . They all share the first appearance of neuropodial bidentate hooded hooks in chaetiger 11 with S. filicornis ( Maciolek 1990, Sikorski 2001). Spio readi differs from S. filicornis in having a narrow peristomium, a distinctly different pattern of pigmentation (paired patches of reddish brown pigment on peristomium and some anterior chaetigers in S. readi , pattern in S. filicornis as above) and a greater number of neuropodial hooded hooks (9–10 in S. readi and 5–9 in S. filicornis ). Moreover, ventral glands or white dots are not described for this species.
Differences between S. malmgreni and S. filicornis are slight; important characters almost match or are unknown from the species description of S. malmgreni ( Sikorski 2001) . In S. malmgreni the inferior neuropodial capillaries first appear on chaetiger 3 or 4 instead of chaetiger 1 as in S. filicornis ; branchiae are absent on last four to seven chaetigers rather than being absent on last six to ten chaetigers as in S. filicornis . The posterior extension of the dorsal ciliated organs also differs, chaetiger 14 at maximum in S. malmgreni and usually chaetiger 11 in S. filicornis . The number of neuropodial hooded hooks is greater in S. malmgreni than in S. filicornis (up to 14 in specimens from Far East opposed to 9 at maximum). Ventral subepidermal glands or any white dots were not mentioned for S. malmgreni . The pattern of pigmentation as well as the length and shape of the nuchal organs are similar. The description of S. malmgreni has to be enhanced and the validity of this species has to be verified as soon as type material becomes available. Differences between S. filicornis and S. arndti sp. nov., are mentioned in the Remarks for the latter.
Intraspecific variations in S. filicornis include the posterior extension of the metameric dorsal ciliated organ, the number and the start of hooded hooks, the intensity of pigmentation and the length of the first branchiae. Some of these characters are obviously size-dependent. The metameric dorsal ciliated organs are present to chaetiger 8 in specimens with 41 and 51 chaetigers, but to chaetiger 12 in specimens with 59 and 60 chaetigers. In some specimens the dorsal ciliated organs do not extend over the full distance between the anterior and the posterior tcb of two subsequent chaetigers. In this case they are restricted to the region of the anterior tcb. This condition may apply to either one (the right or the left) or to both sides of the dorsum. A linear growth of these organs is presumed based on this observation.
In small specimens with 23 chaetigers neuropodial hooded hooks first appear on chaetiger 9, and in specimens with 28 chaetigers in chaetiger 11, implying a loss of anterior hooks shortly after metamorphosis. The number of hooded hooks ranges from 5 in smaller to 9 in the largest specimens. The length of the first pair of branchiae varies between half as long as the second pair of branchiae and the same length (the latter only rarely being the case). Some specimens were only slightly pigmented but the characteristic pattern mentioned above was still visible.
Applying Article 75 of the International Commission on Zoological Nomenclature (2000), a redescription of Spio filicornis ( O.F. Müller, 1776) that provides comparative comments differentiating it from closely related species of Spio has been presented. Type material does not exist. Müller’s original description (1776) and Fabricius’ remarks ( Fabricius 1780, 1785) discussed in this paper indicate that the specimens we recently collected at the type locality belong to S. filicornis ( O.F. Müller, 1776) . The identity of S. filicornis is fixed with the designation of a neotype that is deposited in the Zoological Collection of the University of Rostock, Germany (ZSRO-P2059). We have brought this matter to the attention of our colleagues dealing with Spio concerned about stabilizing the nomenclature of Spio , and they have indicated their support for this decision.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Spio filicornis ( O . F. Müller, 1776 )
| Bick, Andreas & Bastrop, Ralf 2011 |
Spio filicornis ( O .F. Müller, 1776 )
| Fabricius 1785: 264 |
Nereis filicornis O .F. Müller, 1776 : 218
| Fabricius 1780: 307 |
| Muller 1776: 218 |