
Dendrobeania lichenoides Robertson, 1900
|
publication ID |
https://doi.org/ 10.1080/00222930500415195 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE7B54-FFC3-FFF9-DE82-1BD78986B8A3 |
|
treatment provided by |
Felipe |
|
scientific name |
Dendrobeania lichenoides Robertson, 1900 |
| status |
|
Dendrobeania lichenoides Robertson, 1900 View in CoL
( Figure 6E, F View Figure 6 )
Flustra lichenoides Robertson 1900, p 322 , Plate 20, Figures 7, 7a View Figure 7 , 8 View Figure 8 .
Flustra lichenoides: Robertson 1905, p 291 , Plate 15, Figures 91, 92, Plate 16, Figure 105;
O’Donoghue and O’Donoghue 1923, p 23; 1925, p 100; 1926, p 48.
Dendrobeania lichenoides: Osburn 1950, p 167 View in CoL , Plate 25, Figure 6 View Figure 6 ; McCain and Ross 1974, p 13; Dick and Ross 1986, p 89; 1988, p 51, Plate 4D; Soule et al. 1995, p 104, Plate 34A–C.
Description
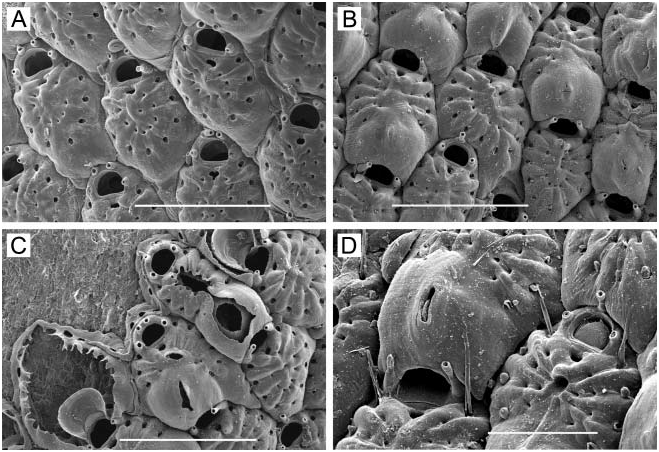
Colony. Recumbent, tan-coloured lobes similar in appearance to some foliose lichens, loosely attached to substrate by radicles ( Figure 6F View Figure 6 ) originating from basal distolateral corner of some zooids; unilaminar, but younger lobes of a colony can overlap older lobes; up to 5–10 cm across.
Zooids. Club-shaped in outline ( Figure 6E View Figure 6 ), rounded distally, widest at orifice, tapering and truncate proximally, each overlapping next distal zooid, 0.60–0.90 mm long (average 50.716 mm, n 515, 3) by 0.23–0.35 mm wide (average 50.306 mm, n 515, 3). Walls weakly calcified, flexible; adjacent zooids loosely appressed to one another; margins sharp. Opesia occupies entire visible frontal surface; proximal gymnocyst covered by preceding zooid; cryptocyst lacking.
Spines ( Figure 6E View Figure 6 ). A straight, erect or distally angled spine at each distolateral corner and a total of three to six more-or-less curved spines along the lateral margins, paired or unequally distributed, angled over the opesia, with no more than four per side.
Avicularia. Lacking.
Ovicell ( Figure 6E View Figure 6 ). Hyperstomial, globose, smooth, imperforate; broader than long, 0.25– 0.33 mm wide (average 50.288 mm, n 515, 3) by 0.18–0.25 mm wide (average 50.202 mm, n 515, 3).
Ancestrula . Not observed.
Remarks
Aggregated colonies formed extensive mats on the undersides of some boulders at East Tongass Narrows.
Distribution
Known from San Francisco ( Robertson 1900, 1905) to Kodiak Island in the Western Gulf of Alaska ( Dick and Ross 1988), with a number of records in between; also recorded from several localities around the Commander Islands ( Grischenko 1997, 2004).
Infraorder ASCOPHORINA Levinsen, 1909 ‘‘Grade’’ ACANTHOSTEGA Levinsen, 1902 Superfamily CRIBRILINOIDEA Hincks, 1879 View in CoL Family CRIBRILINIDAE Hincks, 1879 View in CoL Genus Cribrilina Gray, 1848 View in CoL Cribrilina annulata ( Fabricius, 1780) View in CoL
( Figure 7A, B View Figure 7 )
Cellepora annulata Fabricius 1780, p 436 .
Cribrilina annulata: Robertson 1900, p 234 View in CoL ; O’Donoghue and O’Donoghue 1923, p 30; 1926, p 50; Osburn 1950, p 177, Plate 28, Figure 7 View Figure 7 ; Kluge 1975, p 470, Figure 247; Hayward and Ryland 1998, p 314, Figure 110; Dick and Ross 1986, p 89; 1988, p 51, Plate 4C.
Description
Colony. Encrusting, forming circular or irregular sheets on hard substrates; pink to reddish in colour; largest observed 1.5 cm across, but generally, 0.7 cm; unilaminar, with scattered frontally budded dwarf ovicellate zooids.
Zooids. Autozooids rounded-hexagonal to oval ( Figure 7A, B View Figure 7 ); separated by a groove; 0.35–0.69 mm long (average 50.524 mm, n 515, 3) by 0.30–0.53 mm wide (average50.356, n 515, 3); basal wall uncalcified except for marginal shelf.
Frontal wall. Convex, inflated; frontal shield with 12–17 smooth, rounded, tapering costae connected along their lengths, tips meeting at midline, where there is sometimes a slight keel; each costa with a minute pseudopore at the tip; distalmost pair of costae usually thicker than the rest, forming proximal rim of orifice, the fused tips often raised as a suboral mucro; between adjacent costae are a marginal pore and three to six smaller intercostal pores, their diameter less then the intercostal struts between them; 6–12 intercostal pores in a transverse series across zooid at widest point.
Orifice. Orifice of non-ovicellate zooids 0.09–0.12 mm long (average 50.11 mm, n 515, 3) by 0.14–0.18 mm wide (average 50.160 mm, n 515, 3), rounded distally, broadly concave proximally.
Spines. Usually four (sometimes three) short, stout, erect spines around distal margin of orifice, the middle pair often shorter than lateral pair and sometimes fused as a single broad spine ( Figure 7B View Figure 7 ).
Ovicells. Ovicellate zooids occur in primary layer in some colonies ( Figure 7B View Figure 7 ); smaller than non-ovicellate autozooids, 0.31–0.63 mm long including ovicell (average 50.527 mm, n 515, 2) by 0.21–0.38 mm wide (average 50.301 mm, n 515, 2), with fewer costae (11– 14). Most colonies also have small, frontally budded, dwarf ovicellate zooids ( Figure 7A, B View Figure 7 ) orientated in any direction, 0.20–0.35 mm long including ovicell (average 50.299 mm, n 518, 4) by 0.18–0.25 mm wide (average 50.228 mm, n 518, 4); frontal shield reduced, with three to four short frontal costae in addition to the pair of heavy costae comprising proximal lip of orifice; orifice proximo-distally compressed. Ovicells of primary and dwarf zooids similar, the proximal margin formed by a pair of heavy costae fused at the midline, often ending in a blunt point; distal to thickened costae is a cap-like chamber perforated on top with one to four pores (typically two) that sometimes merge with one another. Pseudopores at tips of proximal ovicellar costae appear as additional small perforations.
Avicularia. Lacking.
Ancestrula . Similar in form to autozooid, though smaller, 0.30–0.38 mm long (average 50.349 mm, n 511) by 0.20–0.30 mm wide (average 50.247 mm, n 511), with four (rarely five) orificial spines. Ancestrula typically first buds a distal zooid, then a pair of distolateral zooids, after which budding continues simultaneously from daughter zooids and proximal half of ancestrula; ultimate compliment of periancestrular zooids is six (one distal, two distolateral, two proximolateral, one proximal, with the proximal zooid abutting the ancestrula, but produced by and connecting to the two proximolateral zooids). Variations occasionally occur in initial budding order.
Remarks
Ketchikan specimens show some differences from material described from other parts of the range of this broadly distributed nominal species. For example, there is a tendency for the middle pair of orificial spines to fuse into a single broad spine ( Figure 7B View Figure 7 ), whereas this has not been reported for material from Britain ( Hayward and Ryland 1998), high latitudes ( Kluge 1975), or Kodiak ( Dick and Ross 1988). In Ketchikan specimens, as in material treated by Kluge (1975) and Dick and Ross (1988), a pair of heavy costal spines forms the thick proximal margin of the ovicell, whereas in British material, the homologous pair of spines curve across the frontal surface of the ovicell, forming a transverse rib separate from the thin proximal margin. Finally, the ancestrular budding pattern seen at Ketchikan seems to differ from that of some other populations. For example, Nikulina (2002) illustrated and noted five periancestrular zooids for specimens from the White Sea; the proximal zooid seen in our material is apparently lacking. These kinds of variation perhaps indicate that across its range, nominal C. annulata may comprise several cryptic biological species; however, there is no basis at present for considering the Ketchikan material as distinct from C. annulata (Fabricius) .
We observed colonies of two different reproductive modes at Ketchikan. In the more common mode, the primary layer lacks ovicellate zooids; dwarf zooids are produced early in colony astogeny and tend to be clustered in the centre of small colonies. In the alternative mode, ovicellate zooids occur in the primary layer; dwarf zooids are not produced early on, and if they occur at all, are in more peripheral positions. We were unable to distinguish any other morphological differences between colonies of the two modes.
Distribution
This is considered a circumpolar, arctic-boreal species. Along the Pacific coast of North America, it has previously been reported from Point Barrow, Kodiak, Cordova, and Yakutat, Alaska ( Robertson 1900; Osburn 1950; Dick and Ross 1988), as well as British Columbia ( O’Donoghue and O’Donoghue 1923, 1926). On the Asian side, it has been reported from as far south as southern Hokkaido Island in Japan ( Mawatari and Mawatari 1981), and the northwest coast of the Sea of Japan ( Kubanin 1997).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dendrobeania lichenoides Robertson, 1900
| Dick, Matthew H., Grischenko, Andrei V. & Mawatari, Shunsuke F. 2005 |
Dendrobeania lichenoides:
| Soule DF & Soule JD & Chaney HW 1995: 104 |
| Dick MH & Ross JRP 1986: 89 |
| McCain KW & Ross JRP 1974: 13 |
| Osburn RC 1950: 167 |
Flustra lichenoides:
| Robertson A 1905: 291 |
Flustra lichenoides
| Robertson A 1900: 322 |
Cribrilina annulata: Robertson 1900 , p 234
| Hayward PJ & Ryland JS 1998: 314 |
| Dick MH & Ross JRP 1986: 89 |
| Kluge GA 1975: 470 |
| Osburn RC 1950: 177 |
| O'Donoghue CH & O'Donoghue E 1923: 30 |
| Robertson A 1900: 234 |
Cellepora annulata
| Fabricius O 1780: 436 |