
Phasmarhabditis bonaquaense, Nermuť, Jiří, Půža, Vladimír, Mekete, Tesfamariam & Mráček, Zdeněk, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4179.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:D1F7B4DE-B169-44B7-8893-E91D01C21B2B |
|
DOI |
https://doi.org/10.5281/zenodo.5630382 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB87FE-BF6A-FF8C-EAEA-E2380268F92E |
|
treatment provided by |
Plazi |
|
scientific name |
Phasmarhabditis bonaquaense |
| status |
sp. nov. |
Phasmarhabditis bonaquaense n. sp.
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Measurements (NDV strain). See Table 1.
Description (based on NDV strain)
Adults. Six distinct, apparent lips surrounding triangular mouth opening seem to be arranged in three pairs, each lip with labial papilla on top. Ring of four cephalic papillae in sublateral position on lips. Small papilla-like protrusion on each lip, laterally, between labial and cephalic ring. Two lateral lips with prominent amphidial opening. Cuticle smooth, annulated, annuli ca 1 µm wide, lateral fields developed. Stoma ca 3.0–3.5 times as long as wide, consisting of short cheilostom, long cuticularized prostom and mesostom, metastom funnel-like, telostom very short. Pharynx distinctly separated into corpus, isthmus, and basal bulb. Pharynx surrounding base of stoma at level of mesostom. Corpus with apparent median bulb occupying ca 60% of pharynx length. Isthmus long and narrow. Nerve ring approximately at mid-isthmus. Basal bulb valvate, ca 18% of pharynx length. Thin and weakly sclerotized excretory pore located slightly posterior to nerve ring, never observed anterior to nerve ring. Amphimictic, both sexes very frequent, displaying approximately equal sex ratio.
Female. Body obese, almost straight when heat killed 2349 ( 1878–2626) µm long. Lateral fields developed, consisting of three prominent central ridges and four incisions. Another indistinct incision are presented laterally (observed only in SEM). Vulval opening located at mid-body region (V% = 54 (45.6–66.7)), vulval lips simple, slightly protruding from body contour and covered with copulation plug in fertilised females. Tail cupola-shaped with long hyaline filiform terminus. Very prominent papilla-like phasmids present, located at base of terminus. Minute post-anal swelling observed. Rectum wide, oblique. Anus: crescent-shaped transverse slit. Genital system didelphic, amphidelphic, ovaries paired, opposed, equally developed and reflexed extending closely to vulva. Spermathecae present, situated at junction of uterus and oviduct, in case of fertilised females filled with sperm cells similar in size and shape to those observed in males. Each uterus ca 316 µm long. Number of eggs in uteri variable, generally ca 24 eggs (range 17–37). Vagina narrow, slightly oblique, with cuticular lining. Matricidal hatching observed very rarely. All females oviparous, ovoviviparous.
Male. Smaller and thinner than female, almost straight when heat-killed, 1829 ( 1414–2121) µm long. Lateral fields developed, with same structure as in females. Bursa copulatrix peloderan, with nine pairs of rays (papillae), 1/1/1/2/1/3, fifth and ninth papillae opened dorsally, other papillae ventrally. One unpaired apparent papilla located on precloacal flap. Prominent papilla-like phasmids near tail tip. Spicules paired, very slightly arcuate, almost straight, with sharp tips. Manubrium slightly elongate. Gubernaculum small, almost straight, only ca 47% of spicule length. Gonad monorchic, extending anteriorly beyond mid-body and reflexing in ca 325 µm from head, testis reflection ca 530 µm. Vas deferens wide, filled with large number of immature sperm cells, ejaculatory duct separated by constriction. Tail long, thick at base and tapering to tip.
Dauer juveniles. Body thin, 902 (808–1050) µm long in heat-relaxed specimens, almost straight in glycerin mounts, slightly tapering towards anterior and posterior ends and mostly ensheathed in second-stage juvenile cuticle. Cuticle annulated in exsheathed juveniles, annuli ca 1 µm wide. Lateral fields beginning posterior to head (from ca 18th annulus), formed by two prominent ridges with wide band (incisure) between them and two marginal incisures. Lateral fields ending at ca 80–90% of tail length. Head smooth, not offset from body contour, six lips, each with one papilla. Outer ring of papillae not observed. Mouth opening closed. Amphidial opening distinct, positioned laterally almost on top of head. Stoma long, 21.1 (19.1–23.9) µm long or ca 8 times longer than broad, forming ca 14% of pharynx length. Weakly sclerotized excretory pore located posteriorly to nerve ring, in front of basal bulb. Metacorpus slightly expanded, isthmus apparent and long, surrounded by nerve ring at half its length. Deirid and hemizonid not observed. Tail long, tapering, with blunt end in exsheathed juveniles. Ensheathed juveniles possessing shorter tail. Hyaline region not developed, anal lips slightly offset from body contour. Phasmids paired, very apparent, located laterally in mid-tail region, ca 1µm in diameter. Rectum long, thin, and oblique, weakly cuticularised.
Molecular characterisation and phylogenetic analysis. This species is further characterised by sequences of ITS, 18S, and 28S regions of the ribosomal DNA.
In all phylogenetic analyses, Phasmarhabditis bonaquaense n. sp. formed a strongly supported monophyletic clade with P. papillosa , P. californica and two strains of an undescribed Phasmarhabditis sp. from Italy. In the 18S tree, a sequence FJ516752 View Materials attributed to Angiostoma dentifera was placed as a sister taxon to P. papillosa and these two sequences show 100% similarity ( Table 2 View TABLE 2 ). This fact strongly indicates that the FJ516752 View Materials sequence in fact belongs to Phasmarhabditis papillosa ( Table 2 View TABLE 2 ).
In the 18S and 28S analysis, several sequences of Angiostoma nematodes fall within the Phasmarhabditis clade ( Figures 5 View FIGURE 5 and 6 View FIGURE 6 ), suggesting paraphyly of the genus Phasmarhabditis . However, further research with a higher taxon sampling is necessary to draw any conclusions about the status of the Phasmarhabditis genus.
Diagnosis and relationships. Phamarhabditis bonaquaense n. sp. shares several characteristics with other species of Phasmarhabditis , such as a smooth, annulated, and longitudinally striated cuticle, a slightly offset head, separated lips, small amphids on the lateral lips, a proximally swollen pharyngeal corpus, paired female genital organs, a midbody vulva, separate and slightly arcuate spicules, peloderan bursa with 9 pairs of rays, large Phasmarhabditis bonaquaense n. sp. differs from P. hermaphrodita , P. neopapillosa , and P. tawfiki mainly in the shape of the female tail. While P. bonaquaense n. sp. has a cupola-shaped tail with a very long filiform tip the others have long, gradually tapering tails. The extremely large phasmids present in the females and males of P. bonaquaense n. sp. are absent in P. hermaphrodita . Moreover, when compared with P. hermaphrodita , the main difference lies in the presence vs absence of males. Dauer juveniles of P. bonaquaense n. sp. further differ from those of P. hermaphrodita in possessing a shorter tail with a blunt tip (85 (66.5–109.5) µm) vs a longer tail with a sharp tip (143 (120–168) µm). Furthermore, the DJs of P. bonaquaense n. sp. are shorter than P. hermaphrodita (902 (808–1050) vs 1097 ( 1016–1140) µm).
The tail shape of the DJs and the large phasmids of adults are among the most apparent differences, also in comparison with P. neopapillosa . Females of P. bonaquaense n. sp. have a significantly shorter tail (85 (66.5– 109.5) µm) than those of P. neopapillosa (157 (141–174) µm). Moreover there are differences in male morphology. Especially, there is an apparent contrast in spicule length (77.1 ± 4.8 vs 68 ± 2.8 µm) and the shape of the manubrium. While the manubrium in P. bonaquaense n. sp. is square-shaped, in P. neopapillosa , it seems to be more triangular and less prominent. The position of the 4th and 5th rays in male bursa seems to be slightly more anterior than in P. neopapillosa . These two pairs of rays were in all observed specimens as being located just in front of or at the level of the cloaca.
The description of P. tawfiki is unfortunately very poor, without any molecular data or photographic material from light or scanning electron microscopy. Despite this complication we can find several differences. The most visible contrast is the tail shape in adult females and probably the bursal formula in a male bursa. While P. bonaquaense n. sp. has nine pairs of rays with a bursal formula of 1/1/1/2/1/3, P. tawfiki , based on the line drawing in the description, seems to have a different formula: 2/1/2/1/3. Furthermore, P. bonaquaense n. sp. has much longer adults and slightly shorter DJs with a significantly shorter pharynx than P. tawfiki . While the males, females, and DJs of P. bonaquaense n. sp. are 1829 ( 1414–2121), 2349 ( 1878 – 2626) and 902 (808 – 1050) µm long, P. tawfiki is 1137 (980–1535), 1716 (1150–2370), 965 (750–1140) µm long. The tail in the females and DJs of P. bonaquaense n. sp. (85 (66.5 –109.5) and 81 (71.6–101.8) µm) is significantly shorter than in P. tawfiki (128 (85– 140) and 144.1 (105–180) µm). In males, the most apparent difference is seen in spicule length, which is longer in P. bonaquaense n. sp (77.1 (68.4–85.9) µm) than in P. tawfiki (63 (54–75) µm).
Phasmarhabditis bonaquaense n. sp. differs from P. californica mostly in the presence of males, in possessing much bigger and more apparent phasmids in females, and in its longer females (2349 ( 1878–2626) vs 1501 ( 1298– 1757) µm). Unfortunately, DJs in P. californica were not described at all.
Males and females of P. bonaquaense n. sp. has a much longer body ( Table 1) and tail (51.1 (46.9–54.7) and 85 (66.5–109.5) µm) than P. papillosa , whose males and females are 1233 ( 1011–1565) and 1590 ( 1202–1932) µm long with tails reaching 43 (28–52) and 106 (73–130) µm. P. bonaquaense n. sp. also has bigger phasmids in comparison to P. papillosa . The most apparent difference can be seen in the male spicules, which are longer and sharper in P. bonaquaense n. sp. than in P. papillosa (77.1 (68.4–85.9 vs 56 (47–67) µm). Similarly, the gubernaculum is also much longer. The manubrium of P. bonaquaense n. sp. seems to be longer and more robust than in P. papillosa (comparison based only on the pictures in the description). The position of the 4th a 5th bursal rays seems to be slightly more anterior than in P. neopapillosa . These two pairs of rays were, in all observed specimens, just in front of or at the level of the cloaca.
Phasmarhabditis bonaquaense n. sp. is a much longer nematode than P. huizhouensis ( Table 1) and the female phasmids also seem to be bigger in P. bonaquaense n. sp. The excretory pore of P. bonaquaense n. sp. is, as is usual in other Phasmarhabditis spp., posterior to the nerve ring. The position of the excretory pore of P. huizhouensis is very unusual, in females it is posterior to the basal bulb and in males anterior to the nerve ring. The most important morphological differences are seen in the size and shape of the spicules. The spicules of P. bonaquaense n. sp. are almost straight with a sharp tip (77.1 (68.4–85.9) µm), while P. huizhouensis has slightly shorter spicules (70.0 (61.1–81.9) µm) that are strongly arcuate with small forked termini. Furthermore, the robust, “rounded” manubrium of P. huizhouensis is wider than the rest of the spicule, while in P. bonaquaense n. sp. the manubrium is much smaller and more gracile.
Type material. Holotype male, paratype males (4 slides with 45 specimens), paratype females (4 slides with 32 specimens) and DJs (4 slides with 64 specimens) were deposited in the Laboratory of Entomopathogenic Nematodes , Institute of Entomology , CAS Biology Centre, České Budějovice, Czech Republic .
± s.đ. (range).
Character Phasmarhabditis bonaquaense n. sp., strain NDV Phasmarhabditis hermaphrodita, Hooper et al. 1999 Phasmarhabditis papillosa, Tandigan De Ley et al. 2016 Holotype Male Female DJs Male Female from Female from DJs from Male Female male slugs bacterial bacterial
culture culture
1 20 20 20 0 20 20 20 12 20 1858 1829 ± 223.6 2349 ± 185.5 902 ± 76.5 Χ 1799 ± 279 1354 ± 115 1097 ± 35.9 1233 ± 184 1590 ± 195 (1414@2121) (1878@2626) (808@1050) (1509@2372) (1186@1525) (1016@1140) (1011@1565) (1202@1932) 18.4 20.8 ± 3.9 17.4 ± 2.3 30.7 ± 4.0 Χ 19.5 ± 3.1 15.2 ± 1.6 26.6 ± 1.2 20.8 ± 2.6 19.9 ± 1.7 (14.0@27.7) (13.3@21.7) (21.7@38.4) (13.6@28.9) (12.4@17.9) (24.0@29.6) (15.6@24.4) (16.0@22.7) 7.0 7.7 ± 1.3 8.8 ± 0.8 5.3 ± 0.6 Χ 7.2 ± 1.1 5.9 ± 0.4 6.3 ± 0.2 6.5 ± 0.9 7.1 ± 0.9 (5.9@10.5) (6.9@10.5) (4.6@6.7) (5.9@9.3) (5.1@6.4) (5.9@6.6) (5.5@7.8) (5.4@9.0) 33.9 36.1 ± 5.7 28.2@5.0 11.2 ± 1.5 Χ 15.8 ± 2.8 13.1 ± 0.7 7.7 ± 0.7 29.2 ± 7 15.5 ± 4.3 (25.8@45.2) (20.0@38.0) (8.3@13.8) (13.2@24.0) (11.6@14.3) (6.6@8.8) (23.2@49.5) (10.9@26.3) 1.3 1.4 ± 0.1 1.2 ± 0.1 4.5 ± 0.5 Χ 3.0 ± 0.3 2.9 ± 0.2 5.6 ± 0.6 1.2 ± 0.3 2.8 ± 0.5 (1.2@1.6) (1.0@1.3) (3.8@5.3) (2.4@3.6) (2.4@3.2 (4.5@6.7) (0.6@1.5) (1.9@3.6) @ @ 54.0 ± 6.0 @ Χ 51 ± 2.6 50 ± 1.6 51.7 ± 2.7 @ 49.9 ± 2.5 (45.6@66.7) (48@60) (48@55) (49@61) (40.5@52.8)
. diam. 101 90 ± 10.9 136 ± 11.6 29.7 ± 3.4 Χ 94 ± 15.8 90 ± 11.2 41 ± 1.4 61 ± 15.7 81 ± 14.6 (70@101) (121@161) (23.5@39.1) (71@118) (75@106) (38@43) (45@88) (57@116)
Stoma length 19.1 19.6 ± 1.6 20.0 ± 1.4 21.1 ± 1.4 Χ 19 ± 0.8 18 ± 1.3 21.6 ± 1.0 18.4 ± 2.4 19.9 ± 2.4 (17.5@22.3) (17.5@22.3) (19.1@23.9) (17@20) (16@21) (20@23) (16.0@22.2) (16.4@25.2)
Stoma diam 6.4 5.8 ± 0.7 6.5 ± 0.4 2.7 ± 0.6 Χ @ @ @ @ @ (4.8@6.4) (6.4@8.0) (1.6@3.2)
region width 15.9 16.7 ± 1.7 18.5 ± 1.7 9.4 ± 0.7 Χ 18 ± 1.2 18 ± 0.7 10.5 ± 0.5 13.5 ± 2.1 15.9 ± 1.7 (15.6@19.6) (15.6@19.6) (8.0@11.1) (16@20) (17@19) (10@11) (10.8@17.9) (13.4@19.7)
Anterior end to 207 196 ± 12.2 231 ± 12.9 151 ± 10.3 Χ 212 ± 11.0 172 ± 12.5 156 ± 8.7 170 ± 3.0 187 ± 13.7
excretory pore (EP) (164@211) (203@246) (129@172) (192@231) (157@189) (143@172) (168@172) (157@208)
Anterior end to mid– 164 144 ± 10.6 160 ± 7.8 95.7 ± 7.5 Χ @ @ @ @ @
corpus (121@164) (144@176) (86.0@117.3)
Anterior end to nerve 184 172 ± 13.9 202 ± 11.6 123 ± 7.9 Χ 166 ± 7.4 141 ± 6.9 127 ± 3.9 139 ± 15.1 168 ± 6.5 (140@195) (188@215) (110@137) (154@177) (131@154) (123@135) (95@151) (152@178)
Anterior end to 266 240 ± 17.9 269 ± 12.3 171 ± 7.9 Χ @ @ @ @ @
pharynx base (ES) (196@270) (250@297) (156@184)
......continued on the next page TABLE 1. (Continueđ
Character Phasmarhabditis bonaquaense n. sp., strain NDV Phasmarhabditis hermaphrodita, Hooper et al. 1999 Phasmarhabditis papillosa, Tandigan De Ley et al. 2016 Holotype Male Female DJs Male Female from Female from DJs from Male Female male slugs bacterial bacterial
culture culture
Pharynx length 247 219.9 ± 18.4 249 ± 12.7 149 ± 8.0 Χ 251 ± 10.4 231 ± 8.9 175 ± 7.6 @ @ (176.4@252.3) (228@276) (134@ 162) (235@270) (212@242) (158@185)
Corpus length @ @ @ @ Χ 115 ± 5.6 107 ± 5.2 85 ± 3.4 93.2 ± 9.5 116 ± 7.5 (109@126) (96@114) (79@91) (80@109) (102@130)
Corpus width @ @ @ @ Χ @ @ @ @ @
Isthmus length 58.7 57.6 ± 6.1 62.8 ± 6.3 45.1 ± 6.0 Χ 64 ± 2.8 59 ± 3.4 48 ± 4.8 49.8 ± 5.0 58.6 ± 3.7 (46.9@66.5) (54.7@74.3) (39.1@54.7) (58@69) (54@63) (39@54) (43.8@58.3) (45.6@63.4)
Bulb length 39.1 38.8 ± 4.2 46.1 ± 2.9 24.5 ± 2.7 Χ 40 ± 1.4 35 ± 2.2 27 ± 2.3 35.8 ± 5.4 40.1 ± 4.2 (31.3@46.9) (43.0@54.7) (19.1@28.6) (37@42) (31@40) (23@31) (27.0@44.5) (34.1@49.4)
Bulb width 35.2 23.8 ± 2.8 39.9 ± 2.6 18.3 ± 1.8 Χ @ @ @ 30.7 ± 4.6 34.7 ± 3.3 (27.4@35.2) (35.2@43.0) (14.3@20.7) (24.6@39.5) (30.0@41.0)
Tail length (T) 54.7 51.1 ± 3.0 85.3 ± 12.9 81.5 ± 7.8 Χ 114 ± 7.8 104 ± 8.6 143 ± 14.1 43 ± 7.5 106 16.5 (46.9@54.7) (66.5@109.5) (71.6@101.8) (99@129) (82@113) (120@168) (28@52) (73@130)
Anal body diam. 43.0 37.8 ± 2.7 71.7 ± 8.3 18.2 ± 1.8 Χ @ @ @ 37.5 ± 7.5 38.4 ± 3.9
ABD) (31.3@43.0) (58.7@86.0) (15.9@20.7) (30.2@49.5) (31.3@44.5)
Testis reflection 505 531 ± 78.7 @ @ Χ @ @ @ @ @ (383@636)
Spicule length (SL) 70.0 77.1 ± 4.8 @ @ Χ @ @ @ 56 ± 7.1 @ (68.4@85.9) (47@67)
Gubernaculum length 35.2 36.1 ± 3.3 @ @ Χ @ @ @ 27.7 ± 5.2 @
GL) (28.6@39.8) (22.4@37.3)
......continued on the next page Character Phasmarhabditis neopapillosa, Hooper et al. 1999 Phasmarhabditis Phasmarhabditis huizhouensis , Phasmarhabditis tawfiki, Azzam 2003 californica, Tandigan De Huang et al. 2015
Ley et al. 2016
Male Female DJs Male Female Male Female Male Female DJs
Pharynx length 246 ± 11.0 290 ± 8.0 180 ± 7.3 Χ @ 189 ± 12.0 214 ± 16.9 212.4 ± 36.15 244.45 ± 27.68 181.17 ± 28.48 (223@261) (277@303) (168 @187) (169@211) (183@251) (155@285) (178@270) (154@255)
Corpus length 119 ± 4.5 144 ± 10.7 80 ± 4.5 Χ 108 ± 9.0 @ 114 ± 23.6 126 ± 18.4 96.1 ± 21.62 (114 @128) (126@168) (71@85) (83@119) (80@160) (100 @156) (75@155)
Corpus width @ @ @ Χ @ 28.9 ± 2.9 37.9 ± 7.1 @ @ @ (25.2@34.2) (24.2@51.7)
Isthmus length 63 ± 5.2 76 ± 6.8 46 ± 3.5 Χ 42.7 ± 4.1 @ @ 53.4 ± 8.2 64.4 ± 8.3 49.89 ± 5.98 (54@74) (65@85) (39@51) (37.4@49.2) (40@70) (48@80) (40@60)
Bulb length 49 ± 3.2 57 ± 5.8 28 ± 1.6 Χ 45.3 ± 2.3 @ @ 43.9 ± 5.1 47.6 ± 7.3 34.55 ± 4.27 (43@55) (48@62) (26@31) (41.6@49.3) (35@55) (36@60) (27@45)
Bulb width @ @ @ Χ 39.5 ± 2.5 40.9 ± 6.4 53.2 ± 9.6 @ @ @ (35.2@43.5) (27.9@52.7) (39.1@71.1)
length (T) 49 ± 2.8 157 ± 15.3 141 ± 7.7 Χ 83 ± 8.9 47.3 ± 8.1 91 ± 6.5 49 ± 3.9 128 ± 22.7 144.15 ± 29.25 (45@54) (141@174) (131@163) (66@94) (35.5@61.4) (80@105) (44@54) (85@140) (105@180)
Anal body diam. @ @ @ Χ 29.7 ± 4.1 41.0 ± 7.0 57.2 ± 5.9 @
ABD) (22.2@38.6) (30.9@55.2) (42.6@67.1)
Testis reflection @ @ @ Χ @ 300@500 @ @ @ @
Spicule length (SL) 68 ± 2.8 @ @ Χ @ 70.0 ± 5.6 @ 63 ± 8.2 @ @ (60@71) (61.1@81.9) (54@75)
Gubernaculum length 33 ± 1.7 @ @ Χ @ 35.4 ± 3.1 @ 36 ± 3.2 @ @
GL) (31@37) (29.9@41.0) (33@40)
Life cycle. Phasmarhabditis bonaquaense n. sp. was isolated from the body of the dead slug M. tenellus and reared in laboratory culture on freeze-killed slugs of the species A. vulgaris or D. reticulatum . This suggests its ability to live on decaying organic material similarly to other Phasmarhabditis species. All described species of Phasmarhabditis are the facultative parasites of terrestrial slugs or snails and P. bonaquaense n. sp. also lives in association with limacid slugs. This species is able to complete its life-cycle on decaying organic animal matter in several days at 15°C. Originally P. bonaquaense n. sp. was found in the mantle cavity of its host. Because only one specimen of M. tenellus was collected during the screening and other collected slugs ( D. reticulatum , D. agreste Linnaeus 1758 , A. vulgaris and Limax maximus Linnaeus 1758 ) were not infected with this species, prevalence and intensity of infection are unknown.
Etymology. The species name refers to the place of origin, the village of Dobrá Voda, bonaqua in Latin translation.
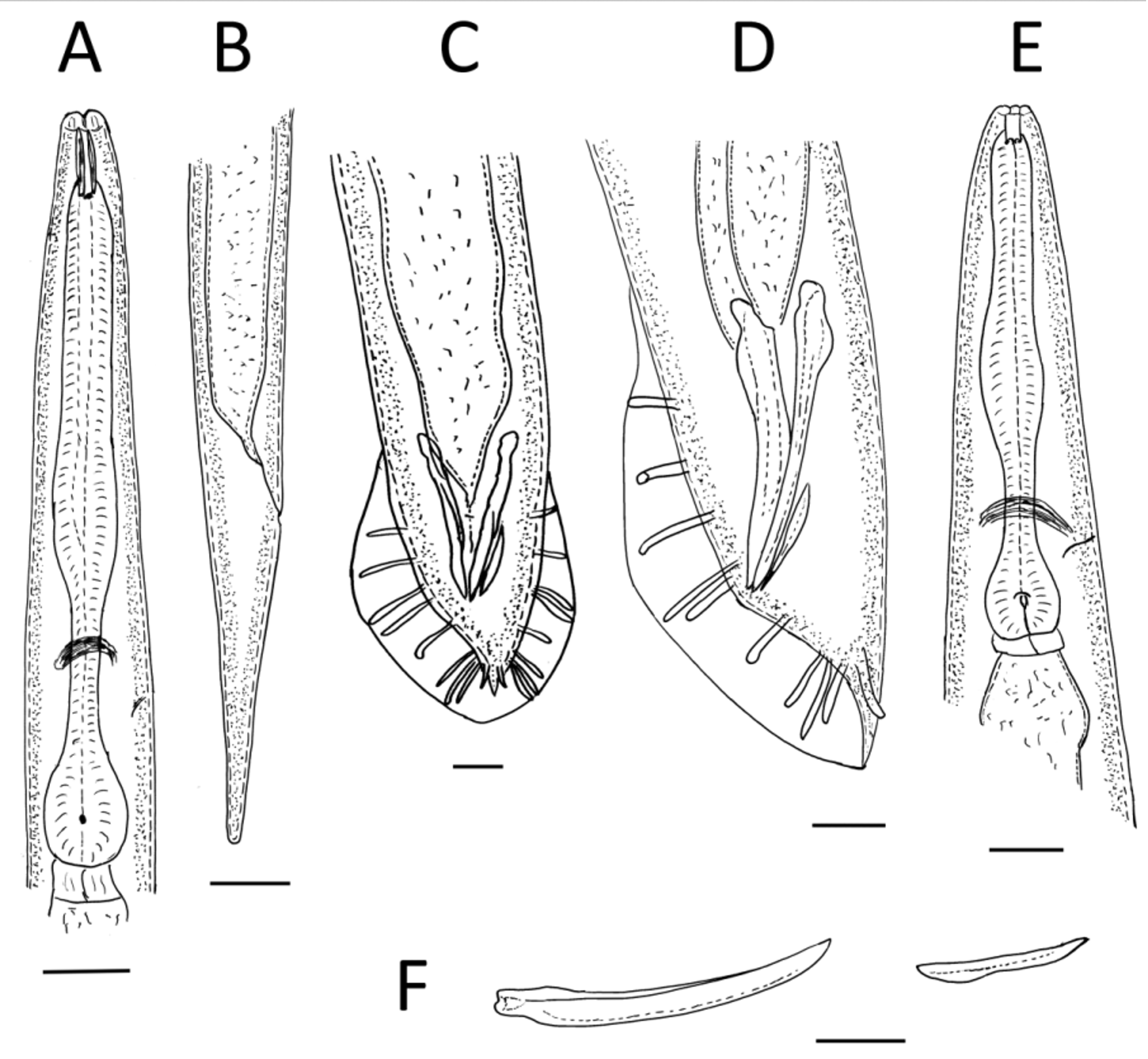
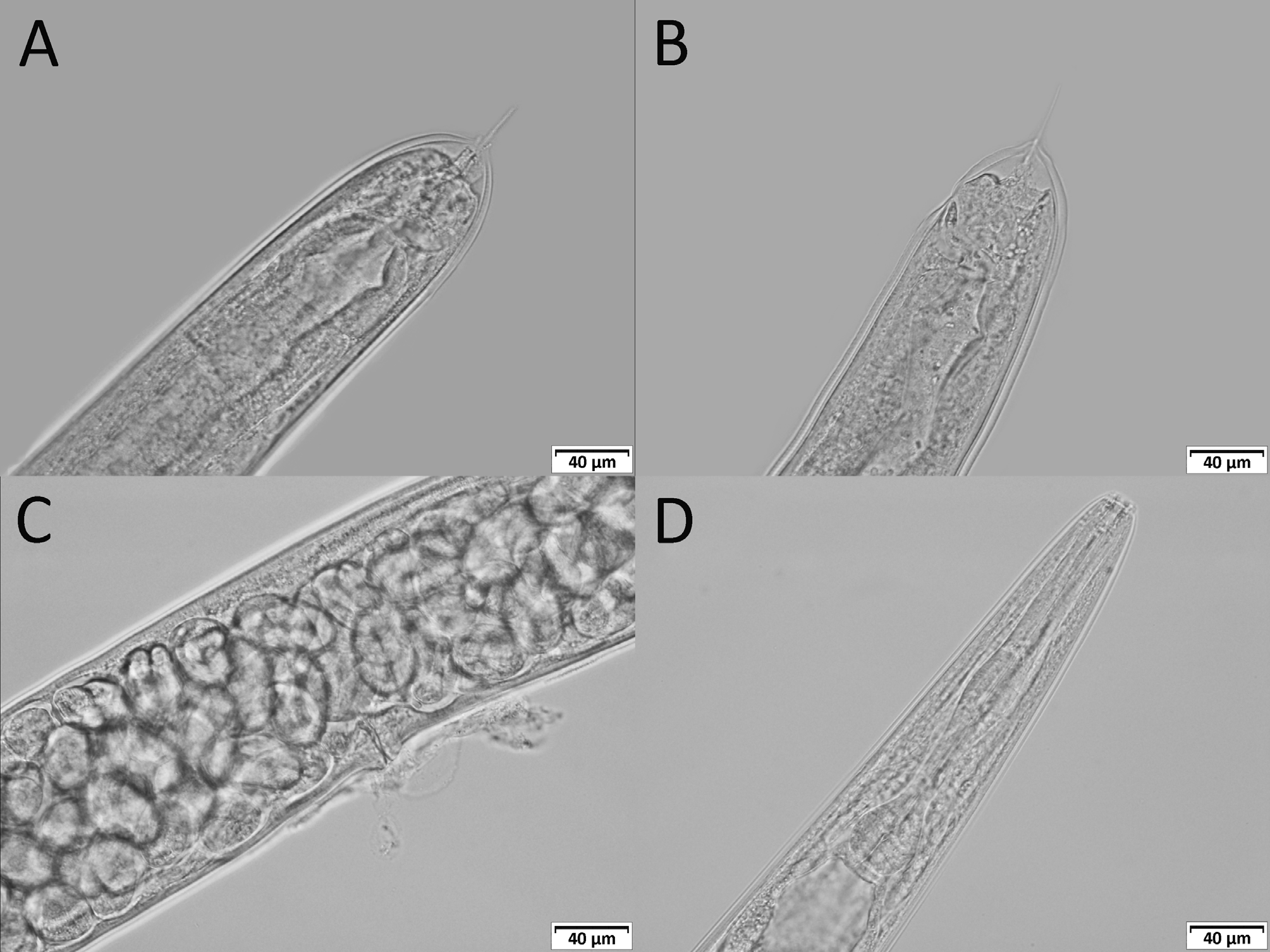




TABLE 2. Pairwise đistances of the SSU region of the rDNA between selecteđ species of Phasmarhabditis anđ other closely relateđ species. Below điagonal: percentage similarity, above điagonal: total character đifferences. Data for P. bonaquaense n. sp. in bolđ.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 ΚΧ017480 Phasmarhabditis bonaquaense n. sp. NDV | 17 | 40 | 10 | 16 | 14 | 50 | 50 | 39 | 38 | 13 | 138 | 40 | 6 | 28 | 155 | |
| 2 KX017477 View Materials Phasmarhabditis sp. GRA | 98.9 | 44 | 12 | 11 | 18 | 52 | 49 | 43 | 44 | 15 | 143 | 46 | 7 | 33 | 163 | |
| 3 KX017478 View Materials Phasmarhabditis sp. CH1 | 97.5 | 97.3 | 34 | 44 | 33 | 37 | 31 | 30 | 30 | 32 | 133 | 24 | 34 | 21 | 151 | |
| 4 KM510210 View Materials Phasmarhabditis californica | 99.4 | 99.3 | 97.9 | 12 | 9 | 44 | 44 | 35 | 39 | 13 | 133 | 37 | 8 | 23 | 155 | |
| 5 KM510211 View Materials Phasmarhabditis papillosa ITD510 | 99.0 | 99.3 | 97.3 | 99.3 | 17 | 54 | 50 | 43 | 42 | 8 | 140 | 43 | 0 | 30 | 157 | |
| 6 EU196008 View Materials Phasmarhabditis sp. EM434 | 99.1 | 98.9 | 97.9 | 99.4 | 98.9 | 45 | 41 | 34 | 35 | 15 | 125 | 33 | 10 | 16 | 157 | |
| 7 KP017252 View Materials Phasmarhabditis huizhouensis | 96.9 | 96.8 | 97.7 | 97.3 | 96.6 | 97.2 | 55 | 42 | 42 | 41 | 152 | 43 | 45 | 37 | 162 | |
| 8 KM510206 View Materials Phasmarhabditis hermaphrodita ITD056 | 96.9 | 97.0 | 98.1 | 97.3 | 96.9 | 97.5 | 96.6 | 22 | 39 | 38 | 140 | 33 | 40 | 27 | 152 | |
| 9 FJ516754 View Materials Phasmarhabditis neopapillosa | 97.5 | 97.2 | 98.1 | 97.8 | 97.2 | 97.8 | 97.3 | 98.6 | 35 | 32 | 136 | 29 | 34 | 18 | 153 | |
| 10 HQ115060 View Materials Phasmarhabditis sp. SA1 | 96.8 | 96.3 | 97.5 | 96.7 | 96.5 | 97.0 | 96.5 | 96.7 | 97.0 | 39 | 114 | 33 | 42 | 14 | 111 | |
| 11 HQ115060 View Materials Phasmarhabditis sp. SA2 | 99.0 | 98.9 | 97.6 | 99.0 | 99.4 | 98.9 | 96.9 | 97.2 | 97.6 | 96.6 | 121 | 34 | 8 | 23 | 123 | |
| 12 EU573705 View Materials Angiostoma limacis | 91.2 | 90.9 | 91.5 | 91.5 | 91.1 | 92.0 | 90.3 | 91.1 | 91.2 | 90.2 | 90.9 | 117 | 127 | 28 | 222 | |
| 13 HQ115062 View Materials Angiostoma margaretae | 97.3 | 96.9 | 98.4 | 97.5 | 97.1 | 97.8 | 97.1 | 97.8 | 98.0 | 97.2 | 97.5 | 91.9 | 41 | 3 | 139 | |
| 14 FJ516752 View Materials Angiostoma dentifera | 99.6 | 99.5 | 97.7 | 99.5 | 100 | 99.3 | 96.9 | 97.3 | 97.6 | 96.5 | 99.4 | 91.1 | 97.2 | 22 | 135 | |
| 15 KC883640 View Materials Angiostoma sp. JLR-2013 | 96.4 | 95.8 | 97.3 | 97.1 | 96.2 | 98.0 | 95.3 | 96.6 | 97.6 | 96.3 | 95.9 | 96.4 | 99.5 | 96.6 | 94 | |
| 16 EU573704 View Materials Agfa flexilis | 90.1 | 89.6 | 90.3 | 90.1 | 90.0 | 90.0 | 89.6 | 90.3 | 90.1 | 90.4 | 90.8 | 85.9 | 90.4 | 90.6 | 88.0 |
| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phasmarhabditis bonaquaense
| Nermuť, Jiří, Půža, Vladimír, Mekete, Tesfamariam & Mráček, Zdeněk 2016 |
Phasmarhabditis papillosa
| Tandigan De Ley et al. 2016 |
Phasmarhabditis papillosa
| Tandigan De Ley et al. 2016 |
Phasmarhabditis hermaphrodita
| Hooper et al. 1999 |
Phasmarhabditis hermaphrodita
| Hooper et al. 1999 |