
Cymbasoma jinigudira, Suárez-Morales, Eduardo & Mckinnon, David, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4102.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9A7BA798-AA7C-4CAA-B42C-1E260CA573E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6091325 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4CA6D-D54F-FFF6-FF12-517E96ED283E |
|
treatment provided by |
Plazi |
|
scientific name |
Cymbasoma jinigudira |
| status |
sp. nov. |
Cymbasoma jinigudira sp. nov.
( Figs 48–51 View FIGURE 48 View FIGURE 49 View FIGURE 50 View FIGURE 51 )
Material examined. Holotype: adult female from North West Cape, Western Australia (21°47.18’ S, 114°7.3’ E), partially dissected, ethanol-preserved; dissected parts mounted on 2 slides in glycerine, sealed with Entellan®. Date of collection: 20th October 1997. Slides deposited in the collection of the Museum of Western Australia (cat. WAM C61308). Allotype adult male from Davies Reef, Queensland, Australia (19°7.340’ S, 146°53.024’ E), partially dissected, ethanol-preserved; dissected parts mounted on 2 slides in glycerine, sealed with Entellan®. Date of collection: 3rd February 1989. Slides deposited in the collection of MTQ, Australia (cat. MTQ W34399).
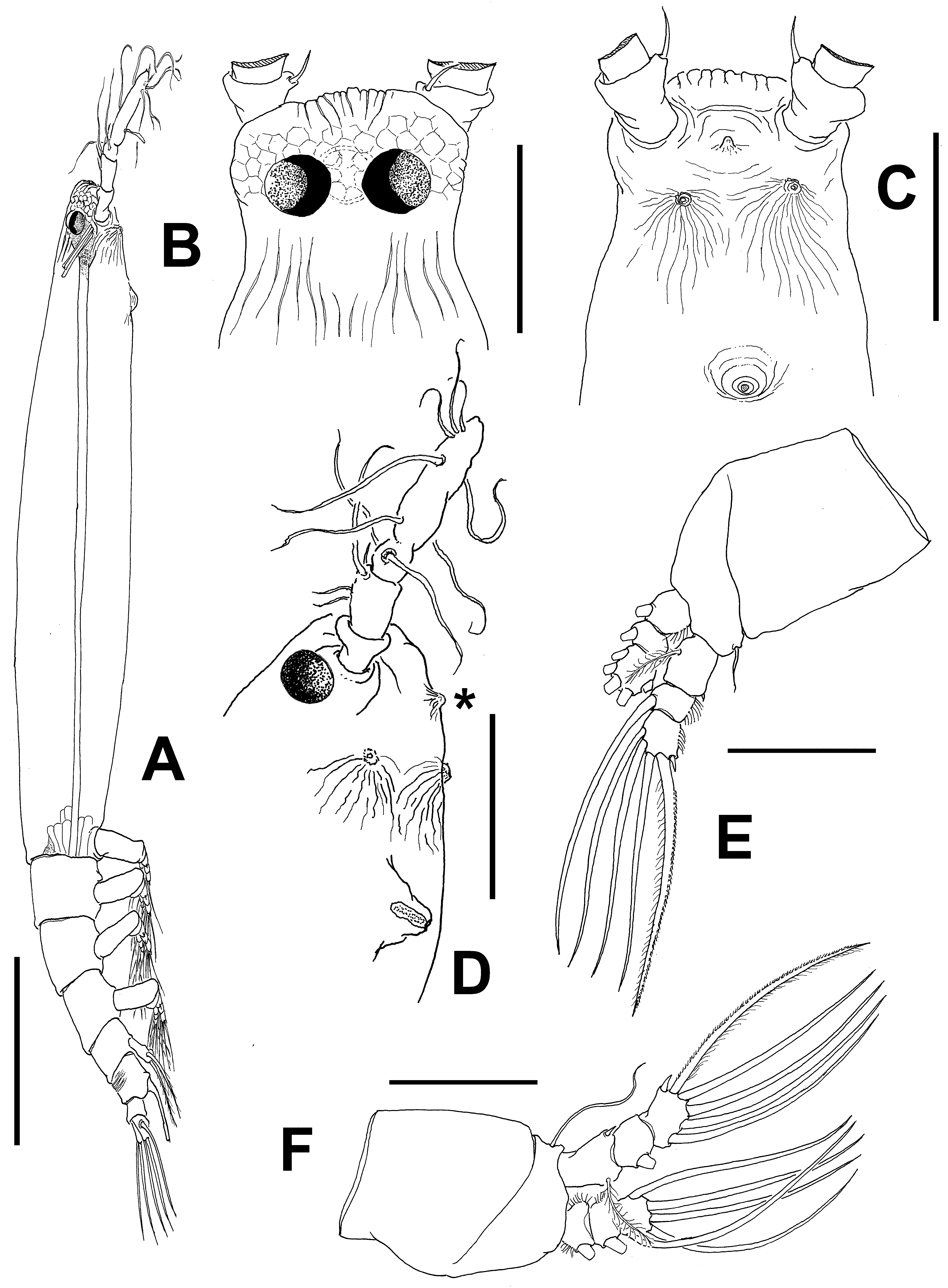
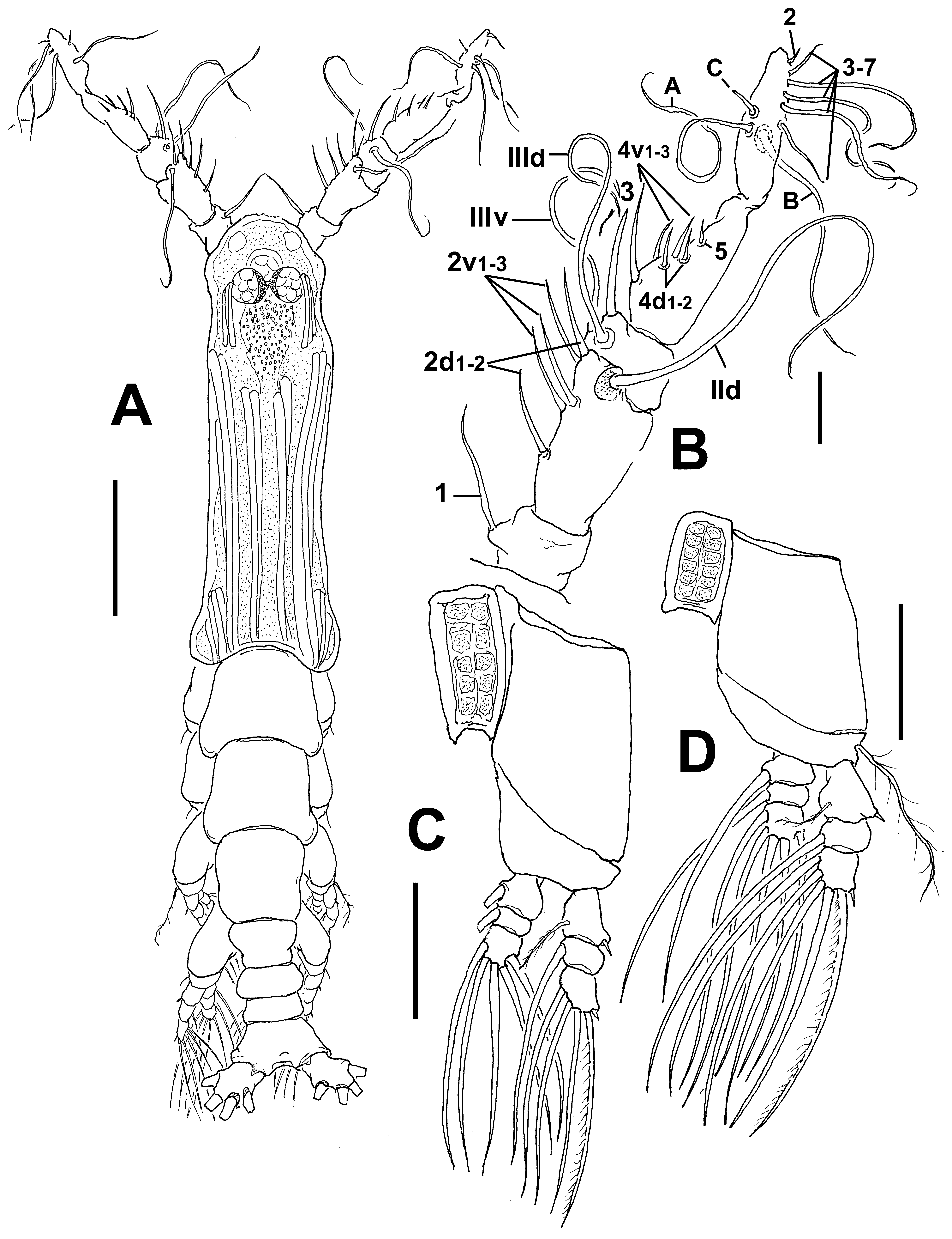
Description of female. Body remarkably elongate, slender ( Fig. 48 View FIGURE 48 A); body length of holotype female 2.82 mm. Cephalothorax approximately 1.95 mm long, representing about 70% of total body length. Midventral oral papilla located at 17% of cephalothorax length. Pair of relatively large ocelli present, pigment cups moderately developed, medially separated, weakly pigmented; ventral cup slightly smaller than lateral cups ( Fig. 48 View FIGURE 48 B). Cephalic area with conspicuous area of deep, longitudinally arranged, cuticular ridges on “forehead” ( Fig. 48 View FIGURE 48 B, C) between the antennulary bases; field of light reticulation overlying ocellar region. Sensillae not observed on frontal area. Additional cephalic cuticular ornamentation including field of longitudinal cuticular wrinkles posterior to reticulation on dorsal surface ( Fig. 48 View FIGURE 48 B). Ventral surface with small medial process (asterisk in Fig. 48 View FIGURE 48 D) and pair of nipple-like cuticular process posterior to antennule bases with extended field of adjacent wrinkles. Other ventral cuticular ornamentation including perioral wrinkles ( Fig. 48 View FIGURE 48 C).
Urosome consisting of fifth pedigerous somite, genital double-somite, anal somite, and caudal ramus, together representing 11% of total body length. Relative lengths of fifth pedigerous somite, genital double-somite, and free urosomite 41.1: 44.4:14.5 = 100, respectively ( Fig. 49 View FIGURE 49 B, C). Fifth pedigerous somite with postero-lateral corners forming rounded protuberances (arrowed in Fig. 49 View FIGURE 49 D). Genital double-somite with dorsal surface bearing transverse pattern of deep cuticular wrinkles on proximal half ( Fig. 49 View FIGURE 49 B, C); with straight ventral margin in lateral view, posterior margin with rounded protuberance (arrowed in Fig. 49 View FIGURE 49 B). Caudal ramus subquadrate, 1.1 times as long as wide, armed with three subequally, long, sparsely setulated caudal setae. Ovigerous spines paired, long, about 1.5 times total body length (2.9 mm), basally conjoined and branch out to form individual spines at level of posterior margin of caudal ramus ( Fig. 49 View FIGURE 49 B). Spines 4.1 mm long, slender, straight at their bases and along shaft, both with distally swollen sections and then tapering apically, one spine slightly shorter ( Fig. 49 View FIGURE 49 E).
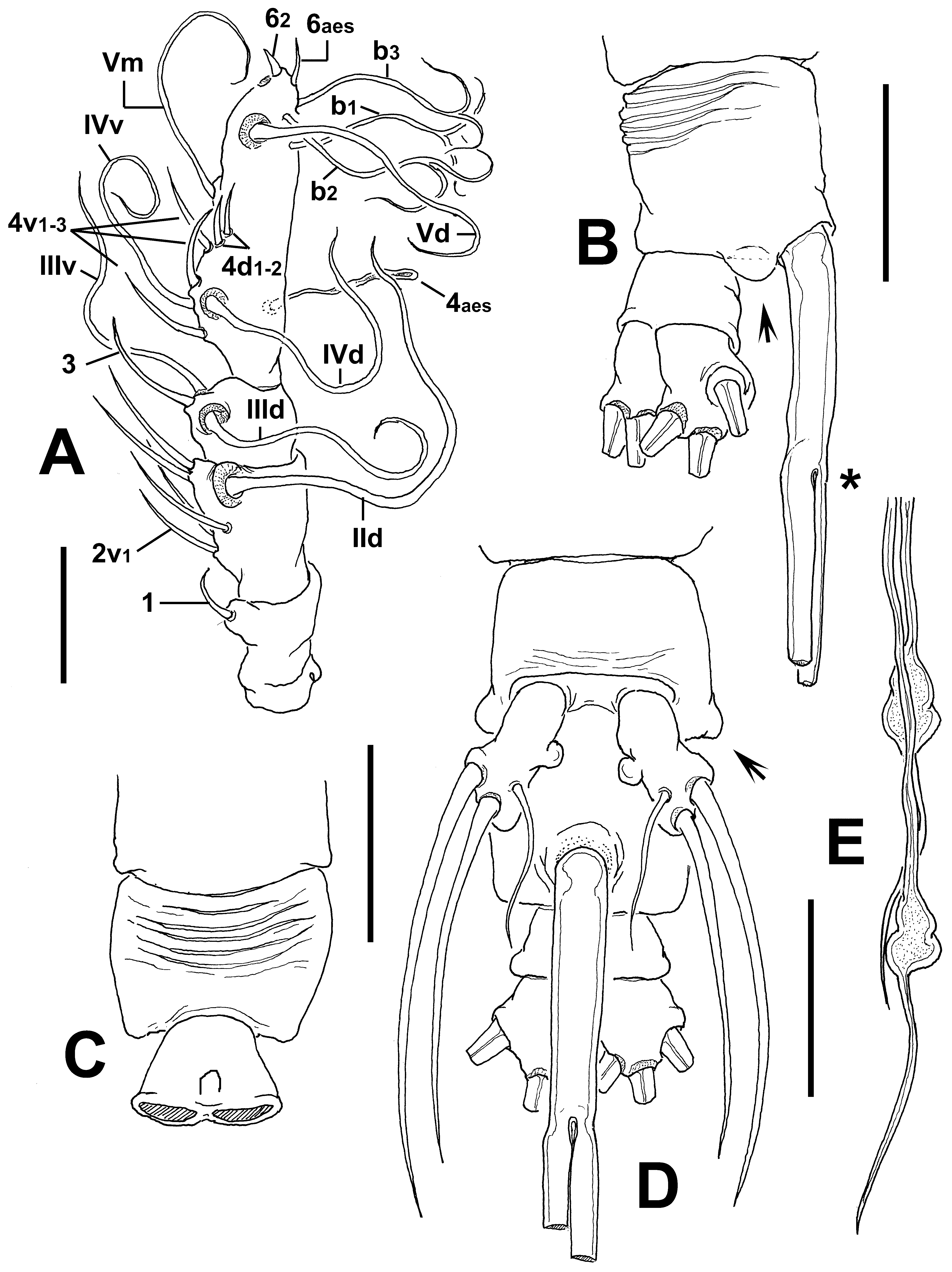
Antennule length 0.46 mm, representing about 17% of total body length and 24 % of cephalothorax length; 4- segmented. Terminal antennulary segment representing 49.6% of antennule length ( Fig. 49 View FIGURE 49 A). In terms of pattern described by Grygier & Ohtsuka (1995) for female monstrilloid antennulary armature, spiniform, curved, slender element 1 present on first segment; elements on second segment 2d1-2, 2v 1-3, these being unusually long, slender, plus element IId. Third segment with long, slightly curved element 3 plus elements IIId and IIIv. Segment 4 bearing elements 4d1,2 and 4 v1-3; elements of 4v group well developed, twice as long as elements of “4d” group. Setae IVd, IVv, Vd, Vv, Vm, and 4aes present. Element 5 absent. Subterminal elements b1-3 dichotomously branched; apical elements 6aes and 62 present ( Fig. 49 View FIGURE 49 A).
Incorporated first pedigerous somite and succeeding three free pedigerous somites each bearing a pair of biramous legs. Pedigerous somites 2–4, together accounting for 22% of total length in dorsal view. Intercoxal sclerites of legs 1–4 subrectangular, widest at base, tapering distally, surface and posterior margin smooth. Bases of legs 1–4 with hair-like lateral seta ( Fig. 48 View FIGURE 48 E, F); on leg 3, this seta about 3.2 times longer, sparsely setulated from proximal half and slightly thicker than those on the other legs ( Fig. 48 View FIGURE 48 F). Endopods and exopods of legs 1–4 triarticulated. Ramal setae all biserially plumose except spiniform outer seta on exopodal segments 1 and 3, and inner seta of first exopodal segment, these latter being relatively long and sparsely setulated ( Fig. 48 View FIGURE 48 E, F). Outermost apical exopodal setae of legs 1–4 with inner margin setulose, outer margin spinulose.
Armature formula of legs 1–4:
Fifth legs medially separated, bilobate. Outer (exopodal) lobe subrectangular, relatively short; armed with three setae, one subdistal, two inserted distally. Innermost seta relatively slender, shortest, about half the length of the other two. Inner (endopodal) lobe reduced, represented by small rounded process arising from middle inner margin of main lobe ( Fig. 49 View FIGURE 49 D).
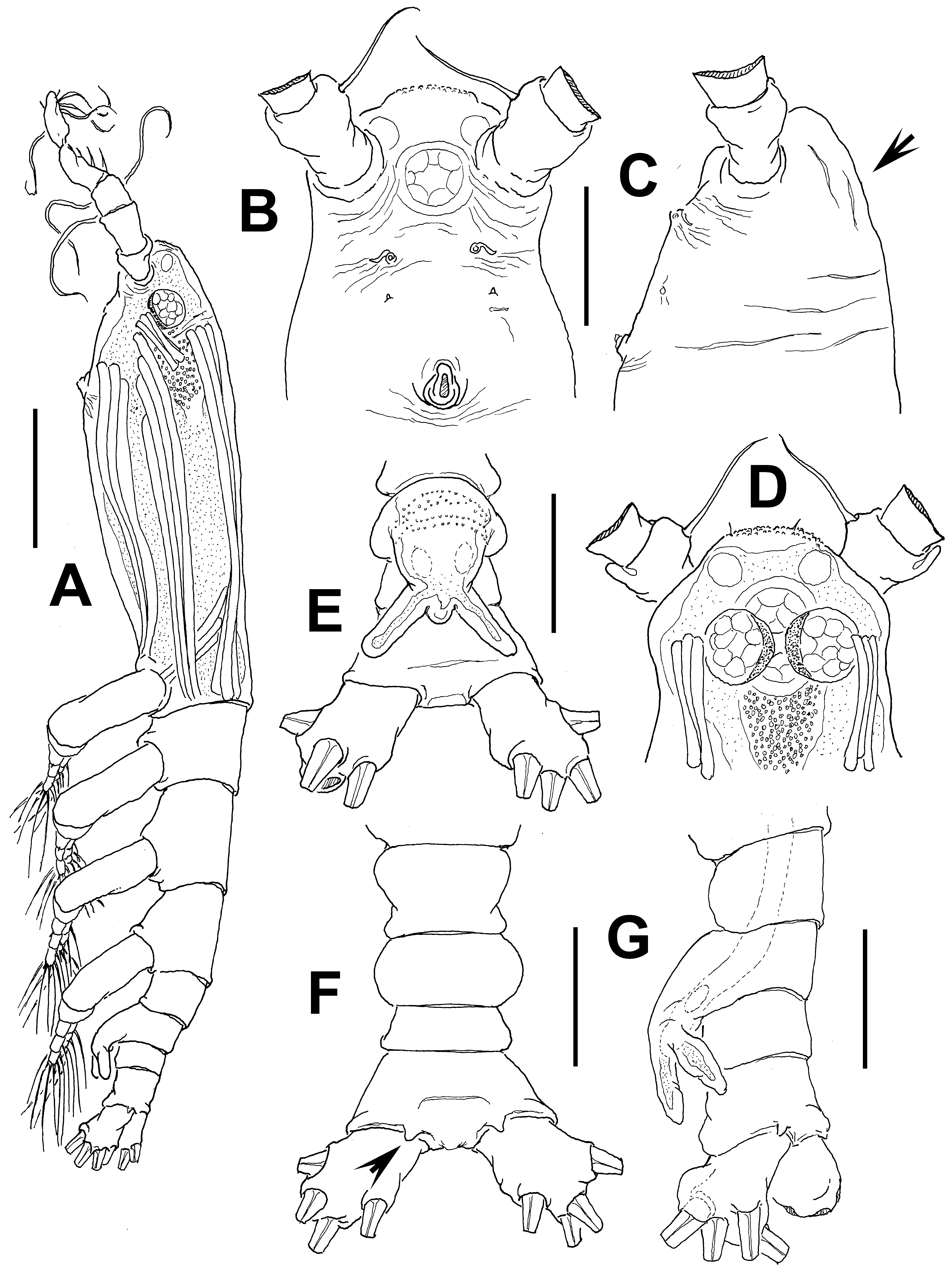
Description of adult male. Total body length 1.24 mm. Cephalothorax 0.61 mm long, representing 50% of total body length ( Figs 50 View FIGURE 50 A, 51A). Midventral oral papilla moderately protuberant, located at 29% of cephalothorax length ( Fig. 50 View FIGURE 50 A–C). Cephalic region slightly protuberant bilaterally in dorsal view, with smooth dorsal surface except for few longitudinal wrinkles (arrowed in Fig. 50 View FIGURE 50 C). Ventral surface between nipple-like processes and post-oral region with few wrinkles ( Fig. 50 View FIGURE 50 B). Pair of dorsal ocelli present, well developed; pigment cups medium-sized. Ocelli medially conjoined, strongly pigmented at inner section. Ventral ocellus slightly larger than eyes ( Fig. 50 View FIGURE 50 D). Pair of frontal sensilla between antennulary bases. Forehead area ornamented with field of minute cuticular papillae ( Fig. 50 View FIGURE 50 B, D). Other cuticular processes on ventral surface include: 1) nipple-like processes with adjacent transverse striae; 2) pair of minute cuticular papilla-like processes posterior to nipple-like processes.
Urosome consisting of fifth pedigerous somite, genital somite (carrying genital complex), preanal somite, and anal somite. Fifth pedigerous somite with smooth ventral and dorsal surfaces, with expanded anterior half ( Fig. 50 View FIGURE 50 F). Genital somite clearly shorter than fifth pedigerous somite, with protuberant, rounded lateral margins; dorsal surface smooth, with rows of papilla-like cuticular processes in anterior ventral half near origin of genital complex ( Fig. 50 View FIGURE 50 E). Preanal somite short, dorsal and ventral surfaces smooth. Genital complex of type II ( Suárez-Morales & McKinnon 2014), represented by pair of divergent, digitiform genital lappets, these being symmetrical, straight. Lappets relatively short, reaching halfway down anal somite, inner and outer margins of lappets smooth ( Fig. 50 View FIGURE 50 E). Rounded, protuberant medial process present at common basal joint of lappets. Anal somite longest of urosome, about twice as long as preanal somite in dorsal and lateral views, comprising 34% of urosome length; somite without lateral constriction but with pair of dorsal spiniform processes along posterior margin (arrowed in Fig. 50 View FIGURE 50 F). Caudal rami subquadrate, approximately 1.2 times as long as wide, about as long as anal somite. Each ramus with four setae.
Antennulary length 0.44 mm. Antennules relatively long, representing 36% of total body length and 69% of cephalothorax length; 5-segmented, all segments separated, with segment 5 located distal to geniculation ( Fig. 51 View FIGURE 51 B). Length ratio of antennulary segments, from first to fifth 10.9: 29: 10.2: 28.6: 21.3 (= 100). Setal element 1 on first segment slender, setiform, remarkably long, reaching proximal half of second segment. Antennulary elements 2v 1-3, 2d1,2 long, slender; element IId present on second segment. Setal elements IIId, IIIv, and 3 present on third segment, latter element remarkably long, reaching middle margin of fourth segment. Fourth segment with elements 4d1,2, 4v 1– 3 present, element 4v 1 longest of group, about twice as long as other “2v-d” spines; other setal elements on fourth segment absent in specimen. Fifth segment with 5 “b”-group setae (sensu Grygier & Ohtsuka, 1995), elements b2 and b3 dichotomously branched distally; element 61 present in distal position. According to Huys et al. (2007) setal nomenclature of the distal segment, elements A–C present, only element 2 present in apical position. Subdistal elements 3–7 present, unbranched.
Incorporated first pedigerous somite and succeeding three pedigerous somites each bearing well-developed biramous legs. Pedigerous somites 2–4, together accounting for 32.5% of total body length in dorsal view. Legs 1– 4 as in other species of the C. longispinosum species-group ( Üstün et al. 2014), except for surface of coxal plates ornamented with spinulose patches ( Fig. 51 View FIGURE 51 C, D).
Type locality. North West Cape, Western Australia (21°47.180’ S, 114°7.300’ E).
Etymology. The species name, a noun in apposition, refers to the coast-frequenting Jinigudira aboriginal people of the area around North West Cape, Western Australia.
Diagnosis. Cymbasoma of the longispinosus species-group with cephalothorax representing 70% of total body length; midventral oral papilla located at 17% of cephalothorax length. Cephalic region with forehead bearing deep longitudinal corrugations, light reticulation overlying area of eyes. Fifth pedigerous somite with postero-lateral corners forming rounded protuberances. Genital double-somite with dorsal transverse wrinkles in proximal half; ventral margin straight, flat in lateral view; distal margin with rounded protuberance. Ovigerous spines fused at base, 1.3 times as long as body. Fifth leg with outer lobe armed with three setae, innermost noticeably shorter, slender; inner lobe globose, small, arising from inner middle margin of exopodal lobe, unarmed. Male with intensely pigmented eyes, anal somite with dorsal spinous processes on posterior margin, genital complex of Type II, with smooth, divergent genital lappets; rounded medial process at insertion of lappets. Four caudal setae.
Remarks. This species from Australia was assigned to the Cymbasoma longispinosum species-group ( Grygier 1994; Suárez-Morales 2011; Üstün et al. 2014) because of the following combination of characters: 1) a long cephalothorax representing more than 65% of the total body length, 2) a quadrate genital double-somite, with its posterior margin wider than the anterior margin of the anal somite, 3) proximally fused ovigerous spines, and 4) a bilobate fifth leg with three exopodal setae, the innermost being smaller and thinner than the others. The nominal C. longispinosum has been recorded in various geographical areas and at different latitudes including the Mediterranean, Norway, the Southern Pacific, the Eastern Tropical Atlantic, the Red Sea, India, Brazil, and the Black Sea ( Üstün et al. 2014). There is a record of a male of this species from the Australian coast off New South Wales ( Dakin & Colefax 1940). The group is currently known to contain at least five species showing subtle but consistent differences ( Üstün et al. 2014).
The new species, C. jinigudira differs from the other members of this group by a combination of characters; with the peculiar arrangement of the forehead ridges, with longitudinal, parallel deep ridges, only shared with C. sinopense Üstün, Terbiyik & Suárez-Morales, 2014 ( Üstün et al. 2014: fig. 2A). Frontal ornamentation is absent in C. morii ( Grygier 1994: figs. 1C, D) in C. janetae Mageed, 2010 ( Mageed 2010: fig. A), and in C. cf. longispinosum from Brazil ( Leite et al. 2010: fig. 2C). In C. chelemense and C. californiense the frontal ornamentation is represented by a swirl-like pattern or simple transverse striae (cf. Üstün et al. 2014: table 1). This character is probably more informative than previously thought when comparing species in this group. The presence of a distinctive shell-like ventral process between the antennulary bases is unique to C. sinopense ( Üstün et al. 2014: fig. 2B); it is absent in the new species ( Fig. 48 View FIGURE 48 C). In C. jinigudira from Australia, the cephalothorax represents 70% of the total body length. This value is similar to that found in C. morii (66–73%) (Martin Thompson 1973; Grygier 1994), C. janetae (70%) ( Mageed 2010: fig. 1A) and C. sinopense (70%) ( Üstün et al. 2014). The other species of the group have a proportional cephalothorax length lower than 68% ( Üstün et al. 2014, table 1), including the specimen illustrated by Bourne (1890) in the original description of C. longispinosum .
The last antennulary segment (46% of the antennule length) and also the relative length of the antennule with respect to the body (less than 15%) are shorter in C. sinopense than in the new species (49% and 17%, respectively). The armature of the fifth leg shows some additional characters for comparison: (1) C. sinopense has the longest inner seta with respect to the length of the outer lobe of the leg (length ratio = 3.6) among the species of the group ( Üstün et al. 2014: fig. 2D, table 1), including the new species (length ratio = 1.1, Fig. 49 View FIGURE 49 D) and (2) C. sinopense has a relatively large, well-developed inner lobe (see Üstün et al. 2014: fig. 2D), whereas this lobe is poorly developed in the new species and is represented by a small rounded lobe arising from the middle inner margin of the outer lobe, as occurs in C. cf. longispinosum from Brazil ( Leite et al. 2010: fig. 2H). The point of bifurcation of the ovigerous spines at the level of the distal margin of the caudal rami ( Fig. 49 View FIGURE 49 B) is a character shared by C. morii , C. chelemense , C. californiense , and C. sinopense but it differs from C. janetae ( Mageed 2010) , C. longispinosum ( Bourne 1890) (original illustrations), and also from C. longispinosum from Norway ( Sars 1921) and C. cf. longispinosum from Brazil ( Leite et al. 2010: fig. 2H). The bifurcation point is located well beyond the distal margin of the caudal rami in the former two species while not reaching its distal margin in the latter two. This character alone would suggest that Sars’ (1921) illustrations of C. longispinosum are not of the same species illustrated by Bourne (1890). In addition, the shape of the fifth leg inner lobe and the length of the innermost exopodal seta are also different. Consequently, it is probable that the specimens from Norway represent an undescribed species of the C. longispinosum species-group. A similar situation has been demonstrated for a species of Monstrillopsis from Norwegian fjords that was identified by Sars (1921) as M. dubia T. Scott, 1904 but in reality represented an undescribed species ( Suárez-Morales & Ivanenko 2004). The point of bifurcation of the ovigerous spines of C. jinigudira appears to display an intermediate condition, being positioned slightly beyond the distal margin of the caudal rami. Also, in the new species the genital double-somite has posterolateral rounded expansions visible in dorsal view and a rounded protuberance at the insertion of the ovigerous spines (visible in lateral position) (arrowed in Fig. 49 View FIGURE 49 B). These processes are absent in the other species of the longispinosum group.
Cymbasoma jinigudira sp. nov. is another of the four species of four Australian Cymbasoma species that possess four caudal setae in the male. It can be distinguished from these congeners by the presence of a conspicuous set of spiniform processes on the posterior margin of the anal somite (dorsal position), which has not been observed in any other male of the genus. The male of the new species lacks an anterior protuberance on the cephalic region, thus diverging from C. bullatum and C. rochai . In C. jinigudira the urosome is relatively short, the genital and anal somites are almost equally long, the anal somite lacks a constriction and is 1.7 times wider than long, thus contrasting with the usual condition (i.e. longer than wide) in the other species of the genus (Suárez- Morales 2000a). The genital lappets are smooth and relatively short, barely reaching the proximal half of the anal somite, and the genital complex is ornamented with rows of papilla-like processes at its base ( Fig. 50 View FIGURE 50 F), a character not previously observed in other species of the genus. The male of the new species closely resembles that of C. tropicum in that 1) the medial margin of the lappet bases has a rounded process, 2) the body proportions (i.e. length of cephalothorax with respect to the urosome, relative length of the antennules) is similar, 3) the caudal ramus has four setae, and 4) the anal somite is wider than long ( Martin Thompson & Easterson 1983). Both species differ in the shape of the body, being clearly more robust in the new species, and in the presence of the spiniform processes on the anal somite. In addition, distinct differences can be discerned in the armature of the antennule. In C. tropicum element 1 (sensu Grygier & Ohtsuka 1995) is short and spiniform ( Martin Thompson & Easterson 1983: fig. 2g) whereas it is remarkably long and setiform in the new species. Also, element 3 on the third segment is short in C. tropicum , reaching the proximal 1/3 of the fourth segment ( Sewell 1949; Martin Thompson & Easterson 1983), whereas it is clearly longer in the new species, reaching well beyond halfway up the fourth segment. There are some differences between the specimens of C. tropicum from the Nicobar Islands and the Maldives ( Sewell 1949) and those from the Arabian Sea ( Martin Thompson & Easterson 1983), particularly in the structure of the genital complex. The Arabian specimens could represent a distinct species.
The previous Australian record of C. longispinosum , based on a male specimen ( Dakin & Colefax 1940, figs. 205Da-d), could be referable to C. jinigudira sp. nov. The structure of the genital complex with a medial rounded protuberance as depicted by Dakin & Colefax (1940: fig. 205Dd) and some additional characters discernible from their illustrations, like the large, wide anal somite and the presence of four caudal setae are reminiscent of the male of C. jinigudira . In addition, all males believed to represent species of the C. longispinosum species group in the literature, based either by their co-occurrence with conspecific females or by explicit designation ( Giesbrecht 1893; Sars 1921; Martin Thompson 1973; Leite et al. 2010) have four caudal setae and a similar genital complex, so these characters inclined us to attribute Dakin & Colefax’s (1940) male specimen to C. jinigudira despite the fact that it was collected at the opposite side of the country. Pending further investigations it is here tentatively assigned to this species.
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |