
Elachistocleis haroi, Pereyra, Laura C., Akmentins, Mauricio S., Laufer, Gabriel & Vaira, Marcos, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3694.6.2 |
|
publication LSID |
lsid:zoobank.org:pub:B1F92852-9EDA-46DB-9662-EFEBD4F67FCA |
|
DOI |
https://doi.org/10.5281/zenodo.5624439 |
|
persistent identifier |
https://treatment.plazi.org/id/03BC87EE-2B73-FFAD-FF20-CC62FCD5F6BA |
|
treatment provided by |
Plazi |
|
scientific name |
Elachistocleis haroi |
| status |
sp. nov. |
Elachistocleis haroi View in CoL sp. nov.
Holotype. FML 24900 adult male collected at El Algarrobal, Jujuy province, Argentina (24°11'27.30" S, 65°13'54.41"W datum WGS 84; 1211 m.a.s.l.), on February 25, 2010, by LCP and MSA.
Paratypes. FML 24894 to 24899, FML 24901 to 24906 and MCN-UNSa 1572 from the type locality; MCN- UNSa 1571 (2 specimens) from Rosario de la Frontera, Salta province, Argentina.
Diagnosis. Last three vertebrae longer than wide in adults was proposed as putative synapomorphy for genus Elachistocleis (de Sá et al. 2012). Presence of this morphological character support the placement of the new species described herein as member of the genus (see osteology description below). Elachistocleis haroi sp. nov. is a small sized species (SVL 25-29.6 mm in males; 26.7-27.7 mm in females), characterized by head length slightly smaller than head width, HL about 84.4% of HW. Dorsum coloration is grayish brown mottled with a paravertebral symmetric pattern constituted by dark spots resembling a pine tree. A mid-dorsal longitudinal bright yellow stripe from the interorbital region, surpassing the post-cephalic transverse skin fold, to vent, and in most cases the line fades to snout. A thin regular yellow line is noticeable on the posterior surface of the thighs and tibiae.
Description of the holotype. Adult male ( Fig. 1 View FIGURE 1 ). Morphometric data is presented in Table 2 View TABLE 2 . The body is ovoid in dorsal view. The head is proportionally small and triangular in dorsal view, slightly broader than long. The head length is about 83.6% of the head width, and about 20.3% of the SVL. It has no cranial crests. The canthus rostralis is rounded; loreal region flat. Snout is “sub-elliptical” in dorsal view and “protruding” in profile. Nostrils small, not protuberant, directed anterolaterally, closer to tip of snout than to eye; internarial distance narrow, smaller than the eye to nostril and interorbital distances. Post-commisural gland poorly developed. Eyes are small, in a dorsolateral position. Texture of the dorsal skin and undersurface of the body is smooth. A transverse skin fold is evident across back of the head, bending downwards slightly behind the eyes to the shoulder. Tympanum is concealed; supratympanic fold absent. Lower jaw is short with a truncate trilobed anterior margin. Tongue is subcircular. Choanae large, subcircular, laterally located and widely separated. Vocal slits present. Premaxillary, maxillary, and vomerine odontophores are absent. Vocal sac subgular, not expanded externally.
Forelimbs are moderately robust, without conspicuous details, like tubercles or crests; fingers with rounded tip, relative lengths of fingers 3>4>2>1. Subarticular tubercles simple and rounded, subarticular tubercle formula I(1), II(1), III(2), IV(1). Palmar and supernumerary tubercles are absent. Fingers without webbing or fringes; prepollex not evident and nuptial pads or cornications absent. Inner metacarpal tubercle oval, smaller than the outer, and divided, resembling three palmar tubercles. Prepollex not evident; nuptial pads or asperities absent. Legs short, robust. Thigh length smaller than tibia and foot lengths; thigh length 34.5% of SVL; tibia length 36.5% of SVL. Heels of adpressed limbs slightly in contact. Knee and heel present a transverse skin fold; no tibial or tarsal ridges; an oval inner but no outer metatarsal tubercle; plantar tubercles absent. Tibio-tarsal articulation does not reach the forelimbs. Toes with rounded tips; relative length of toes 4>3>5>2>1, without webbing or fringes; subarticular tubercles simple, rounded to conical and slightly prominent; supernumerary tubercles absent.
Coloration. In life, dorsum grayish brown mottled with scattered dark spots, with a paravertebral symmetric pattern constituted by dark spots resembling a pine tree ( Fig. 2 View FIGURE 2 ). Dorsal gray color of the snout invades the loreal region to the upper lip border. A mid-dorsal longitudinal bright yellow stripe present from the interorbital plane, surpassing the post-cephalic transverse skin fold, to vent, in some cases this line fades to snout. Venter is immaculate cream, becoming bright yellow to snout and towards the sides. Throat with small dark spots ( Fig. 1 View FIGURE 1 ). A thin regular yellow line is noticeable on the posterior surface of the thighs and tibiae ( Fig. 3 View FIGURE 3 ). Diffuse limit between the dorsal and ventral region from the nostrils to the posterior. Iris gold, marbled with black and brown.
Variation of the paratypes. SVL of males ranges from 25 to 29.6 mm, and from 26.7 to 27.7 mm in females. In a few specimens the mid-dorsal stripe can fade to snout, but always surpass the post-cephalic transverse skin fold. Mid-dorsal stripe can be slightly interrupted in some specimens ( Fig. 4 View FIGURE 4 ).
Osteology. Description based on an adult female (MCN-UNSa 1572) ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ). Skull depressed, longer than wide, and without ridges, exostosis or sculpturations. Dermal elements completely ossified; cartilaginous areas are restricted to nasal cartilage, otic capsules, and suspensorium. Sphenoethmoid is extensively ossified consisting of two elements separated dorsally and ventrally; at the antorbital plane it is expanded through the maxillary arch functionally replacing the palatines. Palatines absents. Prootics form the anterior and medial parts of the otic capsules, a prominent apophyses projecting from the anterior portion of prootics delineate the posterior and interior margin of the optic fenestra reaching and overlapping the medial portion of frontoparietals; exoccipitals form the posterior part of the otic capsules. Occipital condyles are broad and pedunculated. Columella has a small ossified shaft; extracolumella is cartilaginous, wide and crescent-shaped. Operculum is well developed, subelliptical; with a tympanic annulus present. Nasals are large, in contact, and covering the sphenoethmoid. A medial prenasal process is present between the alary cartilages; with a maxillary process large, in contact with the maxilla. Frontoparietals are not fused, but almost in contact, slightly superposed to the prootics and the sphenoethmoid, contacting exoccipitals. Frontoparietal fontanella is narrow. Premaxilla and maxilla lack teeth. Pars palatina of premaxilla is well developed, with a palatine process slightly evident; alary process is nearly vertical and protruded. Maxilla is short, truncated posteriorly, not meeting the reduced quadratojugal; pars palatina developed, pars facialis is low, without a defined preorbital process, superposed to anterior ramus of pterygoid. Vomer is edentate, not contacting the sphenoethmoid, reduced to a thin, elongated, and sharply pointed triradiate element at the inner lateral edge of the longitudinally elongated choanas. Parasphenoid is developed with a cultriform process proportionally wide and with parallel margins covering the medial portion of the sphenoethmoid; the anterior margin is wide and bears a Vshaped incision. Parasphenoid alae are wide, short and truncated. Anterior ramus of pterygoid reaches the antorbital plate and superposes the maxilla; pars medialis is slightly differentiated and separated from the otic capsule and parasphenoid; posterior ramus is fused to the small quadratojugal and to the squamosal. Squamosal is triradiate with ventral ramus developed, oriented posteriorly and confluent with the pars medialis of the pterygoid; otic and zygomatic ramus are vestigial. Mentomeckelians are unfused to each other but synchondrotically related to the dentary, which is projected posteriorly in a lateral, cartilaginous epiphysis, as long as the ossified portion.
Vertebral column has eight presacral vertebrae; the first and second vertebrae are slightly imbricated. The last three vertebrae are longer than wide. Transversal processes are short; diapophysis of vertebrae 2, 7, and 8 are anteriorly oriented; diapophysis of vertebrae 3 and 6 are perpendicular; diapophysis of vertebrae 4 and 5 are posteriorly projected. The process of vertebrae 3 has a well defined apophysis anteriorly oriented. Sacral diapophysis has an expanded epiphysis. Ilium is laying on the projections. Sacrum and urostyle have bicondilar articulation.
Hyoid has a cartilaginous hyal plate, hyoglossal sinus is concave and moderately deep; the alary processes with broad base; posterolateral processes are developed and directed backwards and outwards; hyals do not have anterior processes; tyrohyals are ossified and longer than the hyoid plate, with a cartilaginous distal tip. Pectoral girdle have coracoids strong and ossified, separated medially by epicoracoids cartilages; clavicles as short crescentshaped thin shafts, one-half the length of the coracoid resting at distal end of coracoid. Omosternum is absent; procoracoids are separated and divergent at the union with the clavicles. Scapula is short and strong. Cleithrum is well ossified lying at the anterior margin of suprascapular cartilage. Sternum is cartilaginous, strongly broader posteriorly; with semicircular distal portion.
Humerus has laminar humeral ridge developed. Carpus type C of Fabrezi (1992) formed by radiale, ulnare, distal carpal 5-4-3, distal carpal 2, and element Y; prepollex with proximal and distal elements differentiated. Phalangeal formula is 2-2-3-3. Tarsus formed by distal tarsal 3-2, distal tarsal 1; prehallux with proximal and a small distal element. Phalangeal formula is 2-2-3-4-3 with terminal phalanges of hand and feet simple, short, and slightly curved.
Geographical distribution. We examined collected specimens from El Algarrobal in Jujuy province, and form Rosario de la Frontera and Embarcación in Salta province, Argentina (collection acronyms in Appendix 1). We identified individuals of E. haroi sp. nov. from photographic registries obtained through the colleague M. Lepez from a locality at north of Embarcación, departamento General José de San Martín in Salta province, and from Ruta Nacional 82 near the locality of Teniente General Rosendo M. Fraga, Departamento Matacos in Formosa province. There is also a specimen possibly assignable to E. haroi sp. nov., identified as E. bicolor in Gonzales et al. 2006, from Reserva Natural El Corbalán, Departamento Tarija, Bolivia ( Fig. 7 View FIGURE 7 ).
All localities, except the type locality, belong to Dry Chaco eco-region, characterized by a xeric forest consequence of the arid climate. The type locality is in the transition area between Dry Chaco and the humid Yungas Forest eco-region (Lavilla & Heatwole 2010).
Natural history. The entire area where the specimens of E. haroi sp. nov. were found was modified by human activities. The dominant landscape is open grassland for cattle rising, patches of secondary forest, and urbanized areas. The majority of individuals were found in open grasslands, vocalizing in shallow temporary ponds (less than 40 cm depth) with abundant emergent vegetation. Males were calling inside the ponds, clasped to the emergent vegetation only with their heads above the water surface. Amplectant pairs were found floating exposed on the water surface. Egg masses were deposited in a single layer completely exposed at the pond surface. The eggs were black and the capsule has a lenticular aspect.
Etymology. We take pleasure in naming this species after the Argentinean zoologist Professor Dr. José Gustavo Haro, whose fervent passion for zoology is a constant source of inspiration for LCP, MSA, and MV.
Call description. The advertisement call of Elachistocleis haroi sp. nov. (n = 16 calls from four individuals; three vouchered as FML 24900, FML 24894, and FML 24906) consists of a sustained trill formed by a long and loud, high pitched note of 2353 to 3744 milliseconds of duration (mean = 3186 milliseconds), the dominant frequency peak between 4330 to 4783 Hz (mean = 4560 Hz), and a wide frequency band of 3916 to 5152 Hz ( Fig. 8 View FIGURE 8 ). The call is multipulsed (343 to 605 pulses per call) with a rate of 143 to 162 pulses per second (mean = 152.9 pulses/sec). It is characterized by a modulated frequency rise at the first segment, corresponding to the 41 to 61 initial pulses (mean = 51 initial pulses). Calls have an ascendant frequency modulation at the beginning, with the initial frequency oscillating between 2585 to 3103 Hz (mean = 2643.7 Hz) and the frequency at the end of it oscillating between 4512 to 4576 Hz (mean = 4550.6 Hz). The call rate varies between 3.7 and 5.2 calls per minute (mean = 4.4 calls/min) ( Fig. 8 View FIGURE 8 ). Spectral and temporal parameters of advertisement call of E. haroi sp. nov. and the available descriptions of others species of genus Elachistocleis are summarized in the Tables 3 and 4.
Comparisons with other species. The most noticeable characteristic that allows the distinction of Elachistocleis haroi sp. nov from the spotted belly species of Elachistocleis as E. bumbameuboi , E. carvalhoi , E. cesarii , E. erythrogaster , E. magnus , E. ovalis , E. panamensis , E. pearsei , E. piauiensis , E. skotogaster , E. surinamensis , and E. surumu is the ventral coloration (immaculate belly in E. haroi vs. spotted belly in the listed species). The new species also differs in the following combination of characters (characters of the other species in parentheses): dorsum grayish brown with a paravertebral symmetric pattern constituted by dark spots, resembling a pine tree design (uniformly grayish brown dorsum in E. bumbameuboi , E. carvalhoi , E. cesarii , E. magnus and E. ovalis ; grayish with brighter or darker spots in E. erythrogaster , E. pearsei , E. piauiensis , E. skotogaster , E. surinamensis , and E. surumu ), the mid-dorsal yellow stripe surpassing the post-cephalic transverse skin fold, from the interorbital region, to the vent (absent in E. bumbameuboi , E. carvalhoi , E. ovalis , E. piauiensis , E. skotogaster , and E. surumu ; thin black line from snout to the vent in E. surinamensis ), and by the yellow and well defined stripe on the posterior surface of thighs and tibiae (broad and irregular line on the posterior surface of thighs in E. bumbameuboi , E. carvalhoi , E. carvalhoi , E. magnus , E. ovalis , E. piauiensis , E. skotogaster , E. surinamensis , and E. surumu ; absent in E. erythrogaster ).
The new species differs from spotted belly species of Elachistocleis in several spectral and temporal parameters of its advertisement call. The call is shorter than the call of E. erythrogaster and E. pearsei , and is longer than the call of E. cesarii , E. bumbameuboi , E. panamensis and E. piauiensis . It presents lower pulse rate than E. cesarii and E. bumbameuboi , and has a higher pulse rate than E. erythrogaster and E. panamensis . Also, the advertisement call of E. haroi sp. nov. presents a lower dominant frequency than the calls of E. panamensis and populations of E. cf. ovalis from Rio Grande do Sul ( Brazil), and a higher dominant frequency than E. cesarii , E. erythrogaster , E. pearsei , E. skotogaster , and the population of E. cf. ovalis from Puerto Almacén, Bolivia.
The new species is readily distinguished from the other species with immaculate belly, Elachistocleis bicolor , E. helianneae , E. matogrosso , and E. muiraquitan , as follows (characters of the other species in parentheses). For the comparison with E. bicolor we examined vouchered specimens from Santa Fé and Córdoba provinces in Argentina, and specimens from Uruguay and Rio Grande do Sul state in Brazil. Collection acronyms are in Appendix 1. E. haroi sp. nov. is distinguished from specimens of E. bicolor by the dorsum grayish brown with a paravertebral symmetric pattern constituted by dark spots, resembling a pine tree design (uniformly grayish brown dorsum in all populations of E. bicolor ), the mid-dorsal yellow stripe surpassing the post-cephalic transverse skin fold, from the interorbital region, to the vent (in most cases absent; some specimens from Santa Fé presented a white, thin and diffuse line, only reaching the middle of the dorsum), and by the yellow and well defined stripe on the posterior surface of thighs and tibiae (broad and irregular line on the posterior surface of thighs that fades into ventral coloration in the tibiae, the line is orange in the populations of Córdoba and Santa Fé). From E. helianneae by the diffuse limit between the dorsal and ventral region (sharp color limit between the dorsal and ventral regions), by the well defined stripe on the posterior surface of thighs (broad, irregular), for comparison see color pictures of E. helianneae in Lima et al. (2008). Other differences are the dorsal gray color of dorsum of the snout invading the loreal region almost to the upper lip border (loreal region conspicuously white), and the mid-dorsal stripe surpassing the post-cephalic transverse skin fold, from the interorbital region, to the vent (extending from the tip of snout to vent). From E. matogrosso by the longer SVL (23 ± 1 mm in E. matogrosso vs. 27.5 ± 1 mm in E. haroi sp. nov.), dorsum grayish brown mottled with dark spots, resembling a pine tree design (uniformly grayish brown dorsum), mid-dorsal stripe surpassing the post-cephalic transverse skin fold (extending from post-cephalic transverse skin fold, absent in the head), and by the thin and well defined stripe on the posterior surface of thighs (broad and irregular). From E. muiraquitan by the presence of a grayish brown mottled dorsal pattern with dark spots resembling a pine tree design (uniformly dark brown dorsum), mid-dorsal stripe surpassing the post-cephalic transverse skin fold (mid-dorsal stripe from the post-cephalic transverse skin fold absent in the head), diffuse limit between the dorsal and ventral region (limit between the dorsal and ventral regions well defined), and by the thin and well defined stripe on the posterior surface of thighs (broad, irregular).
The new species differs from other immaculate belly Elachistocleis species in several spectral and temporal parameters of its advertisement call. The call is shorter than the call of E. muiraquitan and E. cf. bicolor from Estación Biológica Beni ( Bolivia), and is longer than the call of E. helianneae , E. muiraquitan , and E. cf. bicolor from La Bola ( Bolivia). It presents lower pulse rate than E. muiraquitan , and has a higher pulse rate than E. helianneae and E. cf. bicolor from Estación Biológica Beni ( Bolivia). Also the advertisement call of E. haroi sp. nov. presents a lower dominant frequency than the calls the two populations of E. cf. bicolor of Bolivia, and a higher dominant frequency than E. muiraquitan .
Tadpole description. The specimen illustrated in Fig. 9 View FIGURE 9 was collected at the type locality of El Algarrobal, Jujuy Province, and presented the following measurements in mm: BL = 8.8, TAL = 15, TL = 23.7, IOD = 5.5, MTH = 4.2, TMH = 2.5, and TMW = 1.5. Table 5 View TABLE 5 summarizes the measurements of tadpoles at different developmental stages.
*specimen illustrated in Figure 9 View FIGURE 9 .
Elachistocleis haroi sp. nov. tadpole is small, TL = 20 ± 2 mm, with a depressed body (body width/body height = 1.41), elliptical in dorsal view. Eyes are small, laterally directed, not protruded, and not visible at ventral view. The eyes and the anterior portion of the oral disc are visible in dorsal view. External nares are absent. Snout is rounded in lateral view, quite pointed and clearly showing oral flaps (visible without microscope). The posterior body end follows a slight anterodorsal curve beginning at the vent tube plane. The spiracle is single, ventral and medial, located at the end of the body, related to the vent tube. The vent tube is medial, fused with the ventral fin, laterally displaced ( Fig. 9 View FIGURE 9 A) and posteriorly oriented, ending just after the body terminus. The tail is quite short, TAL about 62 % of TL. The dorsal fin originates at the tail-body junction. Both fins are slightly expanded at the middle of the tail, MTH higher than the body. The tail tip is narrowly rounded.
Larvae at stage Gosner 35 exhibit an oral disc anteriorly positioned, without keratinized structures (LTRF 0/0) or papillae, and with anterior paired semi-circular dermal flaps with papillae-shaped edges ( Fig. 9 View FIGURE 9 B). Those flaps are the continuation of the snout, visible in frontal, lateral, and ventral views. In dorsal view the central margin of the flaps is also visible at snout tip.
Coloration. Brownish grey in preserved specimens. In dorsal view, anterior central part of the body (from the plane of the eyes) is darker than the anterior part. This dark pattern extends to the posterior part of the body, continuing to the tail. In ventral view, anterior body half white with small grey spots. Dorsal coloration pattern continues into the oral flaps. Viscera are only visible at the posterior half of the body, in ventral view. Tail muscle myosepta are only visible at the anterior part of the tail. The tail fins are mostly transparent with diffuse reticulated and dotted pigmentation at the distal dorsal region. In vivo larvae are mostly black with tail fins smoothly pigmented.
Comparisons with other larvae of Elachistocleis . Elachistocleis haroi sp. nov. larvae mostly coincide in external morphology with other known larvae of the genus, E. cesarii (Magalhães et al. 2012) , E. bicolor (Rossa- Feres & Nomura 2006; Williams & Gudynas 1987), E. erythrogaster (Kwet & Di-Bernardo 1998), E. ovalis (Kenny 1969) , E. panamensis (Vera Candioti 2006) , E. pearsei (Lynch 2006) and E. surinamensis (Kenny 1969) . The most notorious difference is the papillae-shaped edges of the oral flaps. E. bicolor (Rossa-Feres & Nomura 2006; Williams & Gudynas 1987) tadpoles have smooth edges at the oral flaps. Only Elachistoceis sp. in San Pablo, Brazil (Rossa-Feres & Nomura 2006), E. panamensis (Vera Candioti 2006) , and E. surinamensis (Kenny 1969) were reported without smooth edges at the oral flaps.
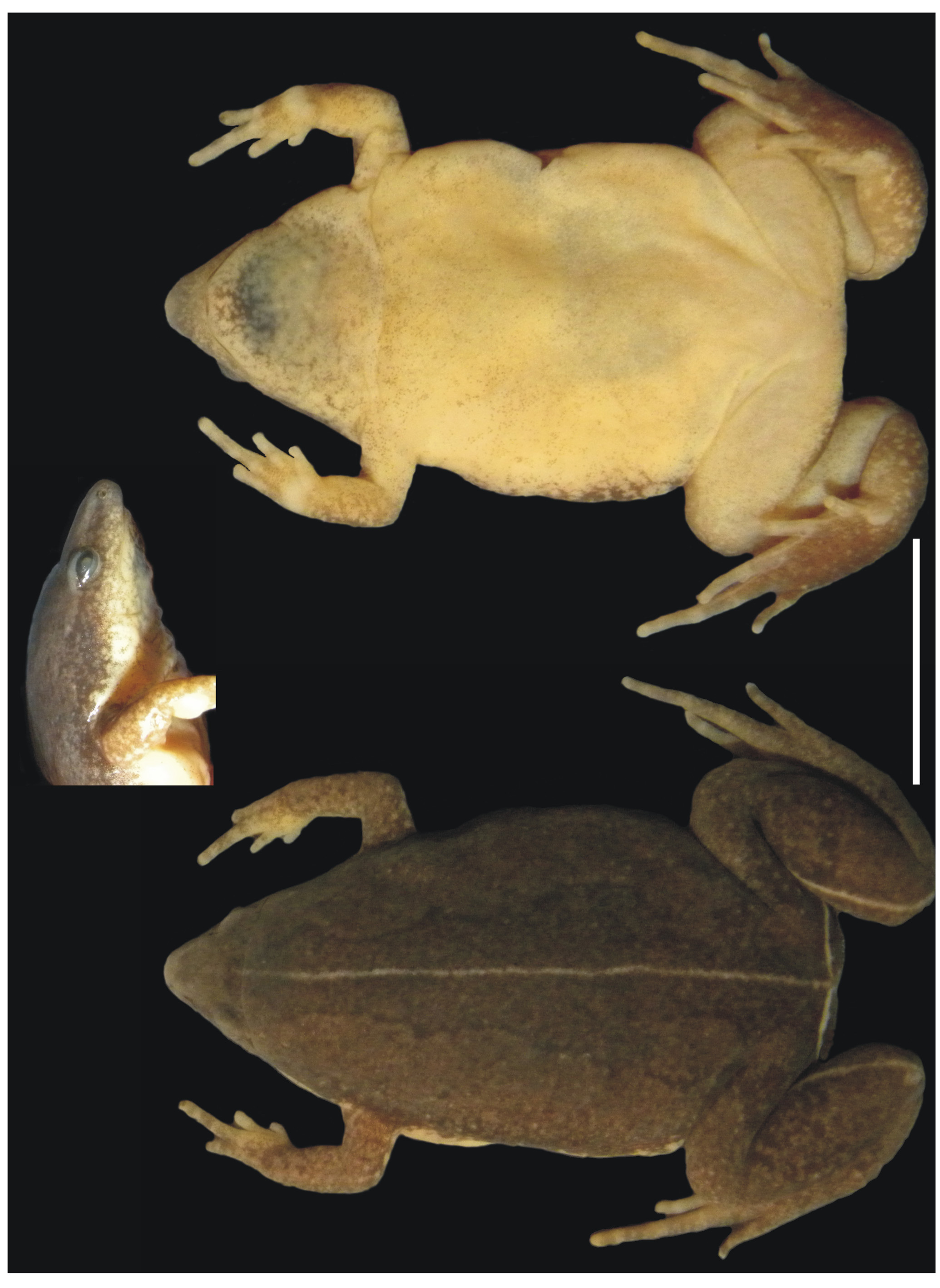



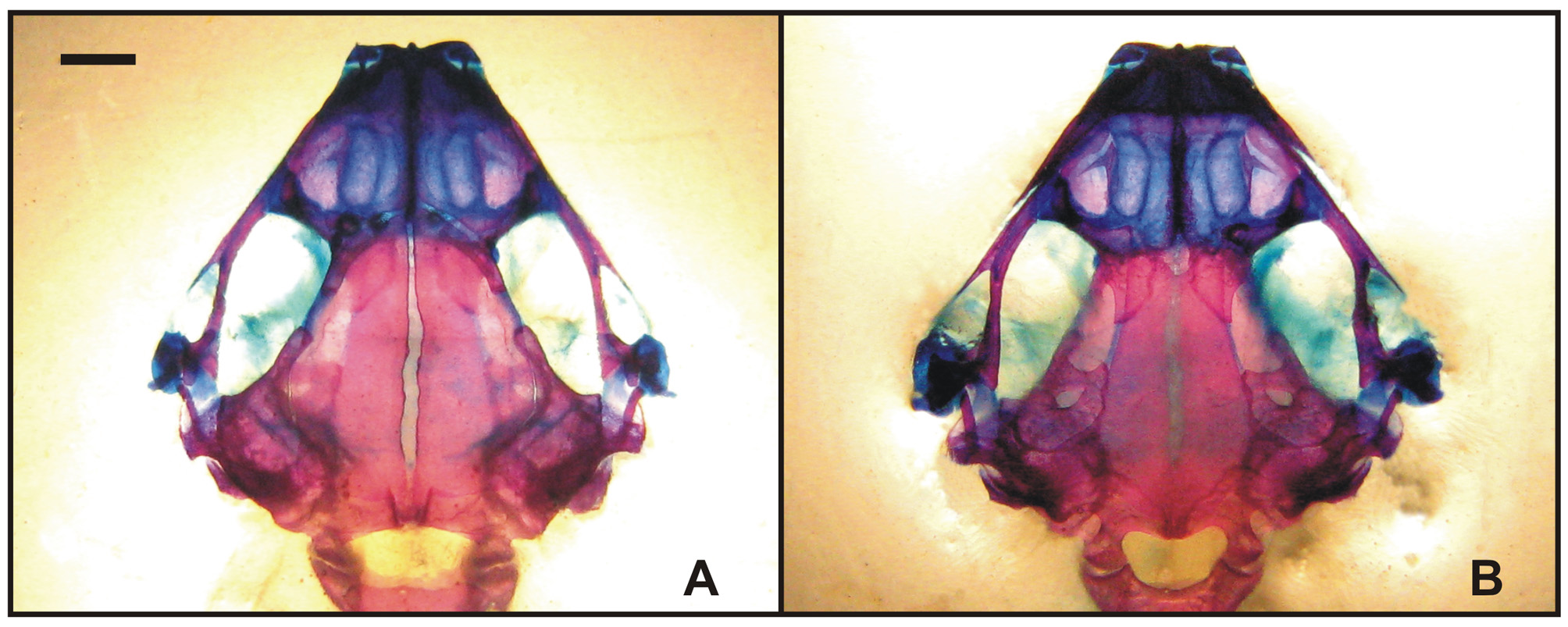
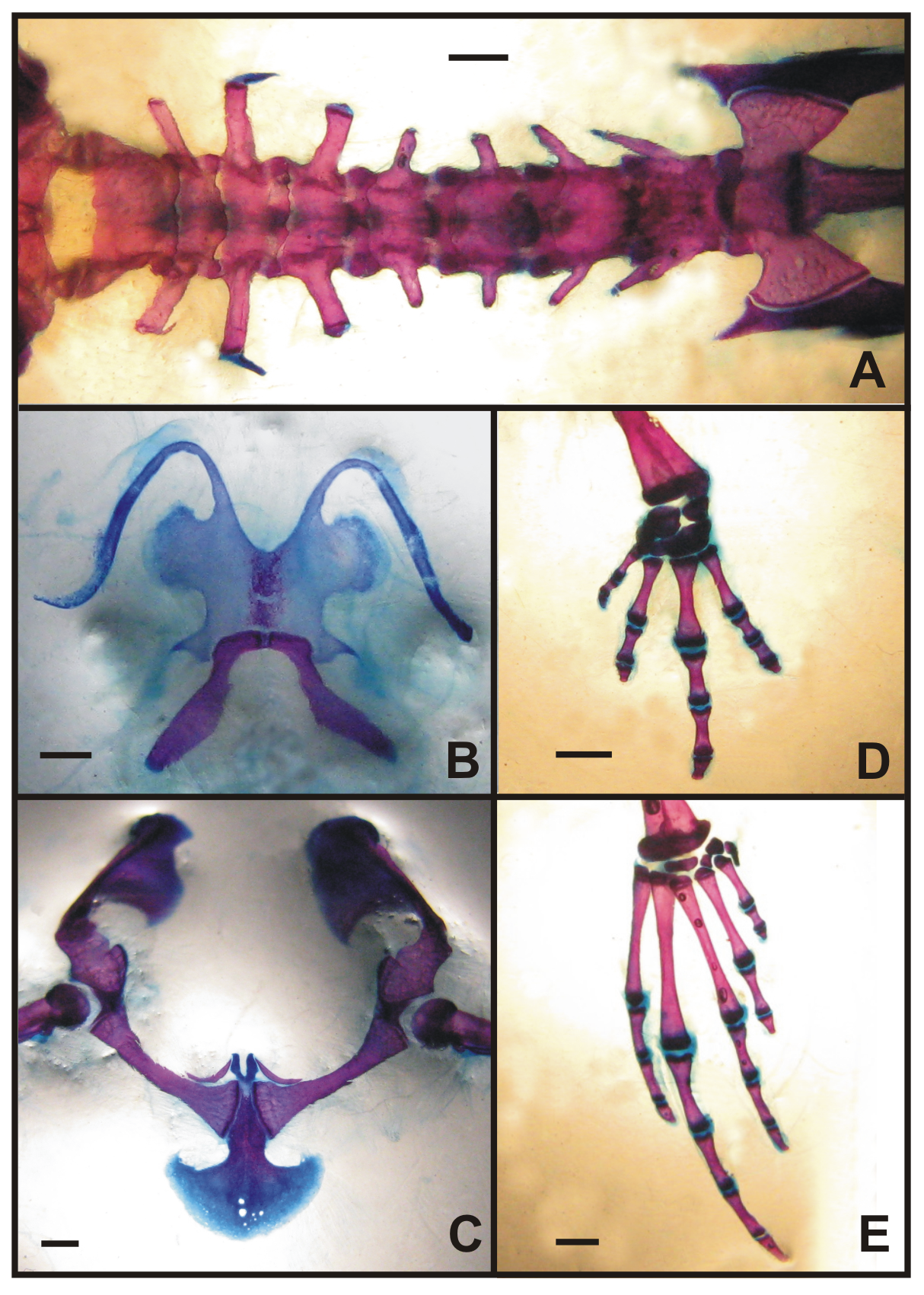
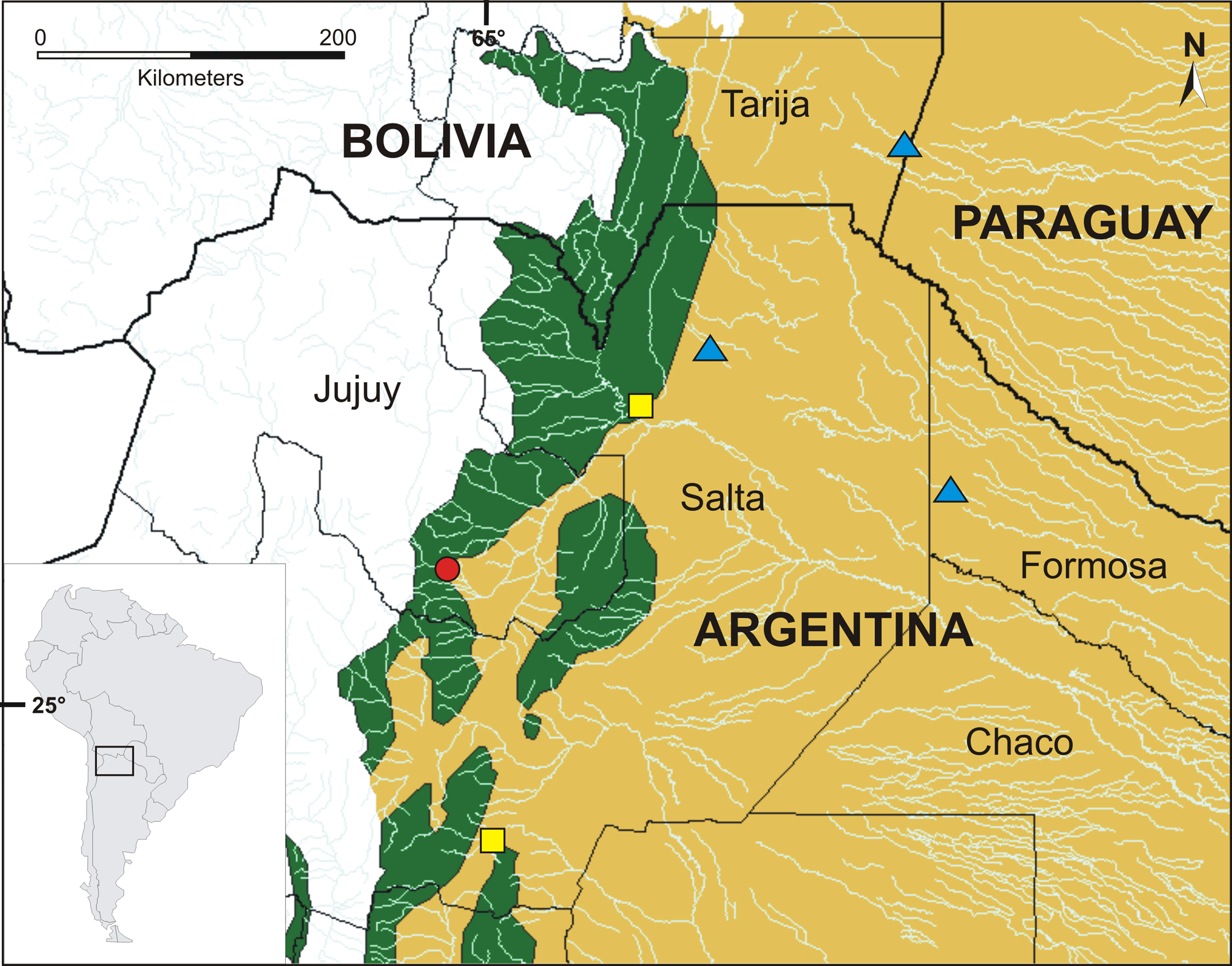
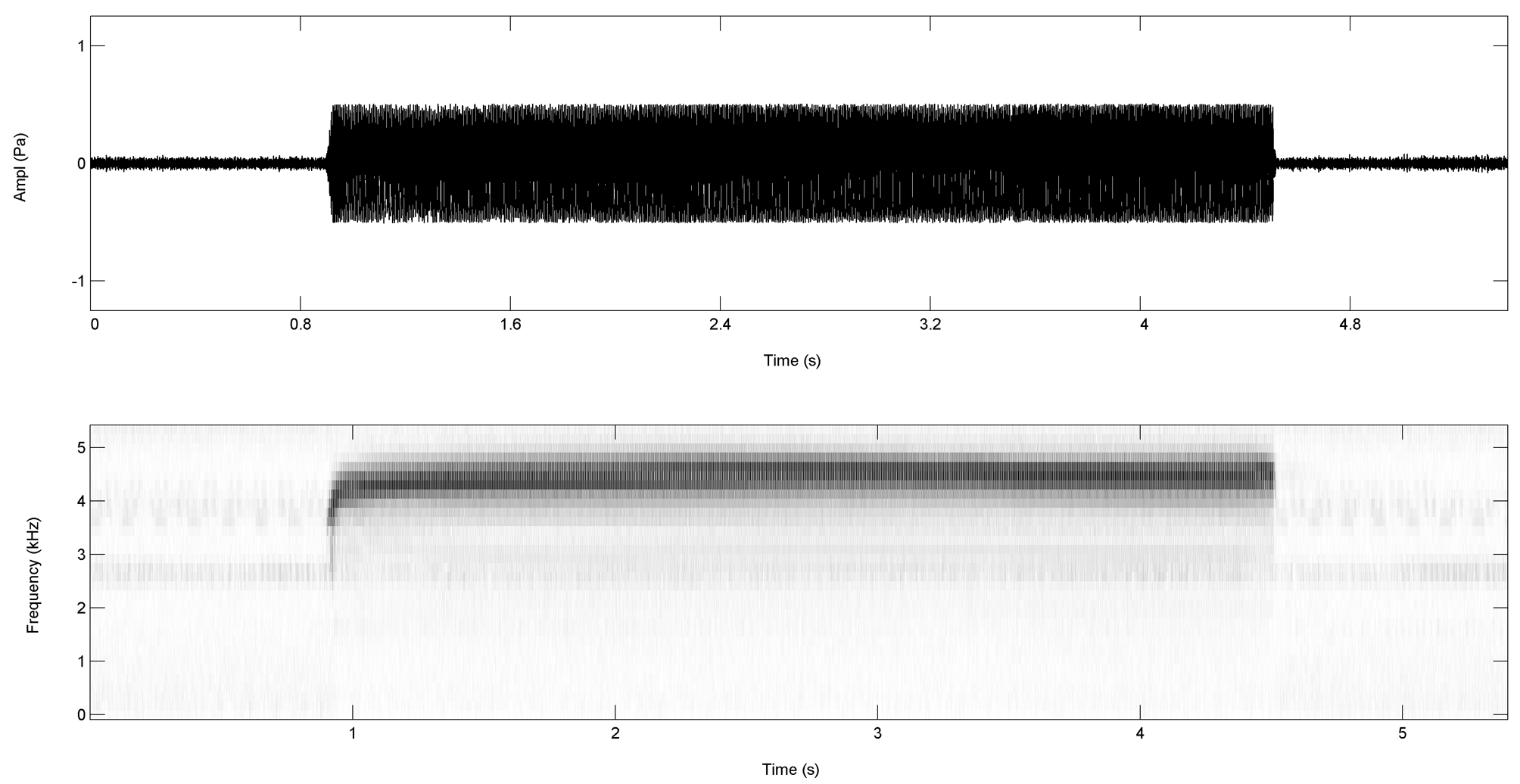

TABLE 2. Measurements: range (mm), mean ± standard deviation of the holotype and paratype specimens of Elachistocleis haroi sp. nov. n = number of specimens. Abbreviations in material and methods section.
| Characters | holotype | males (n = 13) mean | SD Range | females (n = 3) mean | SD Range |
|---|---|---|---|---|---|
| SVL | 27.4 | 27.5 | 1.2 25.1–29.6 | 27.32 | 0.5 26.8–27.8 |
| HL | 5.6 | 5.7 | 0.3 5.1–6.5 | 5.58 | 0.5 5.0–5.9 |
| HW | 6.6 | 6.8 | 0.2 6.5–7.2 | 6.57 | 0.3 6.2–6.9 |
| END | 2.1 | 2.1 | 0.1 2.0–2.3 | 2.13 | 0.1 2.0–2.2 |
| ED | 1.4 | 1.5 | 0.1 1.4–1.5 | 1.43 | 0.1 1.4–1.5 |
| UEW | 0.9 | 1 | 0.1 0.9–1.1 | 0.96 | 0.1 0.9–10.4 |
| IOD | 2.9 | 2.8 | 0.1 2.6–3.1 | 2.68 | 0.1 2.6–2.7 |
| IND | 1.5 | 1.4 | 0.1 1.4–1.5 | 1.38 | 0.1 1.3–1.4 |
| HAL | 5.4 | 5.3 | 0.3 5.0–5.8 | 4.90 | 0.4 4.4–5.2 |
| THL | 9.5 | 9.6 | 0.3 9.1–10.2 | 9.31 | 0.8 8.4–9.8 |
| TL | 10.0 | 10.5 | 0.3 9.9–11.0 | 10.19 | 0.2 9.9–10.4 |
| FL | 11.6 | 11.9 | 0.5 11.1–12.6 | 11.06 | 0.6 10.6–11.7 |
TABLE 5. Measurements (mm) of the ten analyzed tadpoles of Elachistocleis haroi sp. nov. Abbreviations in material and methods section.
| Gosner stage | 35* | 35 | 34 | 34 | 32 | 32 | 32 | 31 | 31 | 30 |
|---|---|---|---|---|---|---|---|---|---|---|
| TL | 23.7 | 22.5 | 20.7 | 23.8 | 19.4 | 17.3 | 22.1 | 18.5 | 17.7 | 18.6 |
| BL | 8.8 | 8.7 | 7.4 | 9.1 | 7.2 | 6.7 | 8.4 | 7.2 | 6.4 | 6.7 |
| TAL | 15.0 | 13.8 | 13.4 | 14.7 | 12.2 | 10.7 | 13.7 | 11.3 | 11.3 | 11.9 |
| MTH | 4.2 | 4.4 | 3.2 | 4.3 | 3.4 | 3.0 | 3.9 | 3.3 | 2.9 | 3.7 |
| IOD | 5.5 | 5.0 | 4.6 | 5.6 | 4.5 | 4.0 | 4.9 | 4.4 | 3.8 | 4.3 |
| TMH | 2.5 | 2.3 | 1.9 | 2.6 | 1.9 | 1.6 | 2.5 | 1.8 | 1.7 | 1.9 |
| TMW | 1.5 | 1.6 | 1.5 | 1.6 | 1.6 | 1.4 | 1.7 | 1.4 | 1.4 | 1.4 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |