
Euura lanatae Malaise, 1921
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4302.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:31B4D326-8D50-41A9-A8A7-69D4427BAD53 |
|
DOI |
https://doi.org/10.5281/zenodo.4901984 |
|
persistent identifier |
https://treatment.plazi.org/id/03B9953B-5C1A-5965-FF48-FB7F239FF864 |
|
treatment provided by |
Plazi |
|
scientific name |
Euura lanatae Malaise, 1921 |
| status |
|
Euura lanatae Malaise, 1921: 105 –106. Described: ♀, ♂, larva, gall, recorded host: Salix lanata . Lectotype, designated below. Type locality: Sweden, Torne Träsk area .
Euura (Gemmura) lanatae: Viitasaari & Vikberg (1985) .
Nematus (Euura) lanatae: Zhelochovtsev (1988) .
Euura (Gemmura) boreoalpina Kopelke, 2001: 197 –201. Described: ♀, ♂, gall, recorded host: Salix glauca . Holotype, ♀, SMF [examined]. Type locality: Norway, Hordaland, Vikafjell near Viksøyri. Preoccupied in Euura by E. boreoalpina ( Lindqvist, 1961) . Syn. nov.
Euura glaucatumida Kopelke in Prous et al. 2014: 53 , replacement name for E. boreoalpina Kopelke.
Notes on types and taxonomy. E. lanatae . Lectotype, ♀, hereby designated: "2", "Torne Tr. Malaise", "Type", " Euura lanatae (ex gall) n. sp." [Malaise's handwriting], "Typus" [red], "NHRS-HEVA000003490", "Lectotype ♀ Euura lanatae Malaise, 1921 des. A. Liston 2016" [red], "DEI-GISHym21299", NHRS. Paralectotypes: 5♀, 4♂, same data as lectotype, with labels "DEI-GISHym21299[–21308]" and “NHRS-HEVA000003491[-3499]”, NHRS . Kopelke (2001) referred in error to the existence of a holotype. The original description of lanatae refers to a type series of 13 females and 8 males. The whereabouts of the missing specimens are unknown.
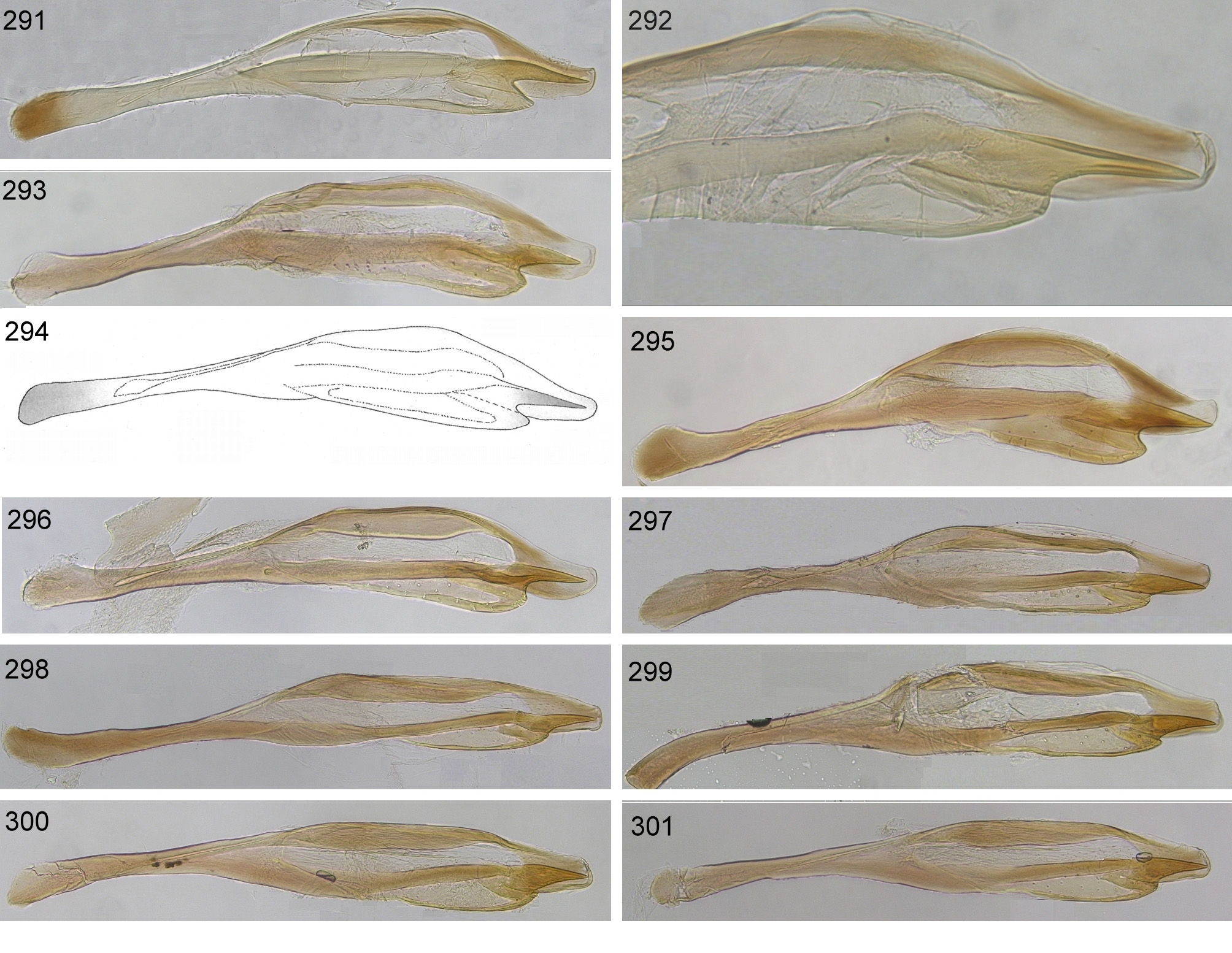
E. glaucatumida is not morphologically distinguishable from E. lanatae . The colour of the tegulae and metafemora, indicated to differ between the two nominal species by Kopelke (2001), is variable in specimens reared from both hosts. On average, specimens reared from S. lanata are larger than those from S. glauca , but this may only be a result of the larger buds of the former host. Kopelke (2001) described and figured the penis valve of E. lanatae as lacking the small notch between the base of the valvispina and the distal lobe of the paravalva, whereas the notch should be present in E. boreoalpina . In the specimens examined, we found this character to display a continuous spectrum of variability ( Figs 297–301 View FIGURES 291 – 301 ), that was not correlated with host plant differences. Specimens of both segregates share a very similar COI barcode, and broader population-genetic analyses have thus far failed to find significant differences between the forms ( Nyman 2002, Leppänen et al. 2014). Accordingly, we regard them as conspecific.
Variability. Female: Body length: 3.3–4.5mm. Clypeus medially black to completely pale. Outer orbit continuously pale to completely black. Tegula white, through brown, to black. Cerci pale to black. Male: 3.4– 4.3mm. Lower inner orbits completely white to completely black. Tegula white, through brown, to black. Total number of specimens examined: ca. 70.
Genetic data. COI barcode not distinguishable from those of E. hastatae , E. lappo , and E. sp. [ Salix myrsinites ].
Similar species. Female; can be confused with E. venusta , but the shape of sawsheath in dorsal view (more evenly tapering in lanatae ), and the frequently dark brown or black tegula (yellow in venusta ) distinguish them. Male; not distinguishable from other nominal taxa in the subgroup.
Bionomics. Host plants: Salix lanata ( Malaise 1921a) , S. glauca ( Kopelke 2001) . Possibly Salix calcicola ( Benson 1962) , if the Canadian specimens referred to are conspecific with E. lanatae . Biology: Malaise (1921a), Kopelke (2001).
Distribution. North Europe (but not British Isles), north to N. Finnmark in Norway; Central Europe (Alps); in Russia probably east at least to the Taymyr Peninsula ( E. mucronata on S. glauca: Roininen & Danell 1997 ). Possibly also North America ( Benson 1962). Occurrence in Sweden: published records; Norrbotten, Lycksele Lappmark, Lule Lappmark, Torne Lappmark ( Malaise 1921a, Haris 2009). Material examined: Härjedalen, Lule Lappmark, Torne Lappmark.
Euura lappo Malaise, 1921a:106 –108. Described: ♀, ♂, larva, gall, recorded host: Salix lapponum . Lectotype, designated below. Type locality: Sweden, Torne Träsk area .
Euura (Gemmura) lappo: Kopelke (1999) .
Notes on types and taxonomy. E. lappo . Lectotype, ♀, hereby designated, NHRS; labels "3", "Torne Tr. Malaise", "Type", " Euura lappo n. sp. Malaise", " Euura saliceti Fall. O. Conde det. 1937", "Typus" [red], " Euura lappo Mal. Kopelke det.", "NHRS-HEVA000003467", "Lectotype ♀ Euura lappo Malaise, 1921 des. A. Liston 2016" [red], "DEI-GISHym21309",. Paralectotypes: 11♀, 6♂, same data as lectotype, with labels "DEI-GISHym21310[– 21326]" and “NHRS-HEVA000003468[–]NHRS-HEVA000003484”, NHRS. Kopelke (2001) referred in error to the existence of a holotype. The original description refers to a type series of 17♀ and 14♂. The whereabouts of the missing specimens are unknown.
Although morphologically very similar to several other nominal taxa in the mucronata subgroup, E. lappo differs biologically from these (except E. lanatae ) in remaining inside the gall to overwinter. Furthermore, Nyman (2002), in an analysis of seven variable enzyme loci, found that E. lappo belonged to a separate lineage from a lineage comprising E. myrsinifoliae and E. hastatae . The validity of E. lappo is also supported by genetic differences in longer mitochondrial COI and nuclear ITS2 sequences (Leppänen et al. 2014).
Variability. Female: Body length: 3.7–4.6mm. Male: 3.1–3.8mm. Female and male: specimens reared by J.-P. Kopelke and E. Heibo from several localities in Norway are much darker than any other bud-galling Euura species so far examined: e.g. metafemora almost entirely black, whereas they are at least apically extensively pale in the other taxa. However, the leg and head coloration of the type specimens is much paler than these Norwegian specimens. Total number of specimens examined: 28.
Genetic data. COI barcode not distinguishable from those of E. hastatae , E. lanatae , and E. sp. [ Salix myrsinites ].
Similar species. Female; externally not distinguishable from others in the subgroup, except laeta , lanatae , and subgemma , or from E. testaceipes and venusta . Male; not distinguishable from other nominal taxa in the subgroup.
Bionomics. Host plants: Salix lapponum ( Malaise 1921a, Kopelke 2001). Biology: Kopelke (2001).
Distribution. North Europe (but not British Isles), north to N. Finnmark in Norway. Possibly also Central Europe ( Czech Republic: Beneš 1968b; galls of E. mucronata on S. lapponum ). Occurrence in Sweden: only known from the Torne Träsk area ( Malaise 1921a).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Euura lanatae Malaise, 1921
| Liston, Andrew D., Heibo, Erik, Prous, Marko, Vårdal, Hege, Nyman, Tommi & Vikberg, Veli 2017 |
Euura (Gemmura) boreoalpina
| Kopelke 2001: 197 |
Euura lappo
| Malaise 1921: 106 |