
Miracarus grootaerti, Wauthy, Georges & Ducarme, Xavier, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.204894 |
|
DOI |
https://doi.org/10.5281/zenodo.6191176 |
|
persistent identifier |
https://treatment.plazi.org/id/03B887F6-FFA7-E10D-FF4E-6D8F0479F85B |
|
treatment provided by |
Plazi |
|
scientific name |
Miracarus grootaerti |
| status |
|
Description of Miracarus grootaerti
( Figs 1–4 View FIGURE 1 )
Diagnosis. Large Miracarus . Prodorsum with rostral slit; anterior free extensions of lamellae ending in large triangular point (or cusp) and abaxially with small tooth; posterior parts of lamellae sloping down and merging into ventral wall of bothridia; with postero-lateral carinae kx extending over bothridia and coming into contact with free border of pteromorphs. Notogaster with lateromedial furrows sl and with lyrifissures appearing each as a small darker spot often well visible surroundig the surface slit. Pleural regions with dorsal carinae kl, subdorsal carinae kb, crook-like carinae kg and parietal carinae kpII; with careniform ribs on acetabular tecta II; with protrusions R and associated dorsal and ventral carinae (e.g. carinae 8 and kt). Ventral region of podosoma with V-shaped carinae cs on mentotectum; with gutter adjacent to circumpedal carinae cir.p; with epimeral furrows s4 well marked laterally. Sejugal stigmata st anterior to pedotecta II. Laterodorsal lobes of vulva with three eugenital setae and setae τ 1 hook-like. Subcapitulum with rutellar microtube prior to lateral lips; with only one pair of adoral setae or. Legs with internal reticulation in several segments; with five setae on femora I; with proral setae on tarsi II, III and IV as very short, triangular spines.
Size, color, sex, cuticle. Mean total length = 321 µm (n = 12, all females; range: 313–334 µm); mean maximum notogasteral width = 224 µm (range: 218–235 µm). Young adults uncoloured or frequently yellowish, suggesting weaker sclerotization; in older individuals, general cuticle chestnut or less often chestnut-brown (yellow in M. abeloosi ). Following elements appearing darker: lamellae, bothridia, carinae tu1 and tu2 of tutoria, pedotecta I and II, protrusions R, discidia, mentotectum; and, among internal elements visible through cuticle in reflected light: apodemes 2, sejugal apodemes, acetabula IV. Rutella, chelicerae and pedipalps colourless or poorly coloured. Thin pore canals belonging to general porosity of cuticle well distinguishable in notogaster in M. grootaerti and lectotype of M. abeloosi , and in M. grootaerti laterally posteriorly to region with cuticular nodules, in ventral part of podosoma and in ventral shield. No male was found among specimens studied in LM in M. grootaerti (in M. abeloosi , samples collected by Lions, 1978: 417, included males and females).
Cerotegument (secretion layer). Studied using SEM and TEM in M. grootaerti . First type of cerotegument consisting of minute (according to some observations, height <0.45 µm and diameter of ca. 0.2 µm at base) and usually spine-shaped protuberances perpendicular to surface of cuticle and with remarkable, dense and rather regular distribution (protuberances frequently arranged in more or less long rows, and cerotegument often also forming short lines mixed with protuberances and rarely making striae on very small surfaces). Detected in following places (frequency not estimated): on dorsal region of prodorsum ventrally to anterior, free extensions of lamellae; in large groove gr of prodorsum up to carinae tu1 of tutoria; ventrally to posterior part of carinae tu2, and both more ventrally and more posteriorly over area equipped with granules and ridges mentioned below (poorly perceptible in Fig. 2 a); dorsally to and ventrally to posterior part of carinae tu3 (not very discernible in Fig. 2 a; dorsally, between tu3 and lower carina associated to parietal carina kpI or in concavity made by lower carina); ventrally to bothridia; in pleural regions dorsally to legs (in Fig. 2 a, posterior limit of pleural cerotegument well visible ventrally to carina kl); on acetabular tecta II and on associated ribs; on and posteriorly to superior wall of pedotecta II; partially on discidia (chelicerae, labrum, ventral side of pteromorphs, some parts of podosoma, and adaxial side of leg segments not examined).
Another type of cerotegument made up of outwardly less high (height frequently <0.15 µm), conical protrusions (pointed or less often truncated) and seen: (1) in some places where distribution of protrusions appears less dense and less regular than distribution demonstrated by protuberances indicated above, as e.g. presumably frequently in genae and in pedipalps (in Fig. 2 b, protrusions are clearly perceptible near infrabuccal slit is and are partially shown on tarsus in right pedipalp; note that cerotegument forms short lines here and there on genae and on pedipalpal segments and, on some pedipalpal tarsi, protrusions and lines appear thicker); another example is given in Fig. 2 c by cerotegument protrusions grouped on tubercle of seta tc’; (2) on borders (as e.g. border of camerostome bl.cam in Fig. 2 b and ventral border of unguis in Fig. 2 c) and on apparent outlines of cuticle (poorly distinguishable in Fig. 2 a except possibly dorsally over short distance in trochanter IV, maybe consisting of foreign materials); and ventrally in tarsus IV in Fig. 2 c); these observations indicate that thin cerotegument possibly covers more or less large part of body and of legs (detailed study not performed; note however that, at higher magnification in Fig. 2 c, very small white points become visible on surface of tarsus). In M. abeloosi, Lions (1978: 405) indicates that no cerotegument exists and that some sort of ‘mucilage’ is sometimes detected on notogaster and on posterior part of opisthosoma (not visible in lectotype of M. abeloosi as well as in M. grootaerti ).
Microsculpture. (not represented in Figs 1 View FIGURE 1 and 3 View FIGURE 3 a). In M. groootaerti , according to observations in LM and SEM, microsculpture consisting of thin granules (diameter ordinarily <1 µm) infrequently mixed with microridges. Appearing particularly well perceptible in following places (frequency not estimated): partly on rostrum; on dorsal surface of lamellae except anterior ending cusp and posterior sloping part of lamellae; laterally on dorsovertex; posteriorly on carinae tu1; on carinae tu2 and tu3; ventrally to carinae tu2 (distinguishable with difficulty in Fig. 2 a; consisting of thin granules anteriorly and, posteriorly, of somewhat larger granules, diameter of ca. 1–1.5 µm, with clearly less dense distribution and mixed with some diversely inclined ridges spreading out, ventrally, to parietal carina kpI and, posteriorly, to pedotectum I); ventrally to carinae tu3; on carina kx; on pteromorphs and on region of notogaster close to pteromorphs; partially on superior wall of pedotecta I (granules well perceptible posteriorly to carina ku in Fig. 2 a); posteriorly to pedotecta I, on and dorsally to zone of acetabular ribs (granules not very numerous and also somewhat larger here); on mentotectum; on legs partially on bulb in trochanters III and IV, on all femora, and on tibiae III and IV.
In lectotype of M. abeloosi , not studied in detail (however, granules fairly discernible in LM on lamellae including both cusps but apparently not on posterior part of lamellae, laterally on dorsovertex, and where microsculpture appears to be lacking in M. grootaerti , as e.g. on large groove gr of prodorsum, as shown in Figs 3 View FIGURE 3 a of Lions, 1978, and on genae). In both species, with nodules (diameter of ca. 1.5–2 µm, sometimes ca. 2.5 µm) in pleural regions dorsally to legs II and III (in M. grootaerti , frequently extending more posteriorly than in Fig. 2 a, yet less markedly than in Fig. 3 View FIGURE 3 a of Lions, 1978, in M. abeloosi ); often appearing well rounded in face view and frequently suitably protruding. Appearing more compact in M. abeloosi ; for example, in lectotype, with 12 nodules at left and 13 at right on surface of 100 µm2 dorsally to processus R and, in M. grootaerti , number of nodules counted on equivalent surface and in similar place between five and 11, n = 6 (in lectotype, nodules laking posteriorly to bothridium, at right, on rectangular area of ca. 140 µm2 and, at left, on less large surface more or less similar to area shown in Fig. 3 View FIGURE 3 a of Lions, 1978).
Prodorsum. ( Fig. 1 View FIGURE 1 ). As in M. abeloosi , dorsophragma absent and lateral margin of camerostome simple, without genal incision. Lengthening of prodorsum clearly less pronounced in M. grootaerti (compare Fig. 1 View FIGURE 1 in this work with Fig. 1 View FIGURE 1 of Lions, 1978). Yet, with regard to notogaster, prodorsum with similar breadth in both species (width of prodorsum measured between anterior borders of bothridia/maximum width of notogaster measured in undissected specimens posteriorly to pteromorphs ratio averaging 0.56, range: 0.55–0.58, n = 6, in M. grootaerti and 0.57 in specimen of M. abeloosi represented in Fig. 1 View FIGURE 1 of Lions, 1978; lectotype not studied). Note that animal in Fig. 1 View FIGURE 1 of Lions (1978) is not orientated according to greatest length (in such orientation, animal is more inclined up), yet appears to be orientated so that median zone of ventral region of podosoma is perpendicular to plane of observation; and, in M. grootaerti , animal with similar orientation in Fig. 1 View FIGURE 1 .
Remarkable traits. Slit of rostral hood (imperfectly shown in Fig. 1 View FIGURE 1 ). Similar to M. abeloosi in shape and size. Sometimes, as in M. abeloosi , with membrane partially obstructing slit: three slits with membrane 4.3, 4.3 and 4.7 µm broad; and, mean breadth of 6.1 µm, range: 4.3–7.7 µm, n = 15, detected in slits without or with reduced membrane. Slits both fairly narrowed (<5.0 µm) and without membrane observed in three specimens.
Anterior, large and free extensions of lamellae. Differing from M. abeloosi in: (1) in dorsal projection, anterior extensions of lamellae partially covering rostral region whereas, in M. abeloosi , rostral region being totally covered by anterior extensions of lamellae (compare Fig. 1 View FIGURE 1 in this work to Fig. 1 View FIGURE 1 of Lions, 1978); (2) anterior ending cusp and abaxial tooth shaped as in M. discrepans , M. hurkai and M. senensis whereas, in M. abeloosi , ending part consisting either of two small and broad cusps located on both sides of lamellar seta le, as seen on right and left lamellae in lectotype, or of only one adaxial cusp, as at right in Fig. 2 a and at left in Fig. 2 b of Lions, 1978; in M. grootaerti , extremity of cusp appearing acuminate in dorsal projection (left side in Fig. 1 View FIGURE 1 ) or acute and wide (not shown) or less frequently truncate and broad (right side in Fig. 1 View FIGURE 1 ), and lateral borders of cusp often symmetrical; and moreover, with tooth usually touching tubercle of seta le (in some instances, tooth absent and seta le then situated more adaxially, i.e. more or less in middle of anterior extension of lamella); (3) both symmetrical adaxial borders divergent anteriorly and parallel posteriorly (both borders occasionally coming into contact or overlapping posteriorly) whereas, in M. abeloosi , both adaxial borders being parallel over long distance ( Fig. 1 View FIGURE 1 of Lions, 1978).
Posterior, sloping downward part of lamellae. In both species, postero-ventral deviation shown by lamellae evidenced, in lateral projection ( Fig. 2 a), by sudden pronounced bend of free border bl and, in dorsal projection ( Fig. 1 View FIGURE 1 and, for M. abeloosi , Fig. 1 View FIGURE 1 of Lions, 1978), by less marked deflection of bl in abaxial direction, and is accompanied by change in orientation of lamellae (according to observations in serial sections, lamellae appearing to rise anterior to deviation and to go down in posterior part; not studied in M. abeloosi ). Ventral side with one (more infrequently two) transverse ridge (not seen in Fig. 1 View FIGURE 1 ) running onto large groove gr and usually seeming to meet posterior stria of groove gr; note that: (i) contiguous to lamellae, grooves are recesses for distal part of legs I, when legs are folded against body, and posterior part of lamellae is probably used to protect genu as in M. abeloosi ( Lions, 1978: 405) ; (ii) both grooves meet together anteriorly ( Fig. 1 View FIGURE 1 ) and, posteriorly, limit of grooves consists of convex wall (only very small part of wall is shown on both sides through cuticle of notogaster in Fig. 1 View FIGURE 1 ) which is united to ventral wall of bothridia (abaxial boundary of convex wall labelled 1 in Fig. 2 a and appearing well marked, careniform in lateral projection). In M. abeloosi , according to lectotype, with six or seven transverse striae running onto gr and ending posteriorly to carina tu2 of tutoria (see below). Note that posterior parts of lamellae cannot be regarded as ‘sublamellae’ observed in other Brachypylina (e.g. Grandjean, 1953a: 120) because arriving at bothridia (cf. Grandjean, 1952: 33).
Postero-lateral carinae kx. As in M. abeloosi , consisting of two parts (posterior portion of anterior part and posterior part being in contact with dorsal wall of bothridium). Anterior part of kx: enlarging as coming near and reaching bothridium ( Fig. 1 View FIGURE 1 ); apparent outline appearing broadwise V-shaped anteriorly and U-shaped posteriorly according to observations in serial cross-sections; generally prolonged anteriorly by slender ridge either running transversally ventrally to free border of posterior part of lamellae (and then touching free border of lamella, as in Fig. 1 View FIGURE 1 at left, or not) or running longitudinally to deviation of lamella (and then arriving at deviation, as in Fig. 1 View FIGURE 1 at right and in Fig. 2 a, or even sometimes going beyond); with comparable ridge on both sides in lectotype of M. abeloosi (yet seeming to be located just posterior to deviation of lamella at left and appearing longer and coming into contact with free border of posterior part of lamella at right). Posterior part of kx: short and clearly going down (hardly seen at left in Fig. 1 View FIGURE 1 , and not shown in Fig. 2 a; though variable, declivity of posterior part always marked); with anterior portion of border of pteromorphs leaning against posterior part of kx.
Rostral hood. (cf. Grandjean, 1952: 33, 1966a: 494). In both species, with incomplete limbus occupying in plane of symmetry less than half-length of rostral hood. With short duct prolonging alveole of setae ro and running inside limbus as seen in M. abeloosi in Figs 2 a and 3a of Lions (1978) and detected on one side in lectotype; no duct found in M. grootaerti . Superior wall with median convexity (two divergent lines close to slit mark partially lateral limits of convexity in Fig. 1 View FIGURE 1 ) bearing laterally rostral setae ro and ending posterior to setae ro (also found in lectotype of M. abeloosi , yet setae ro appearing to be inserted on lateral limits of convexity), and ordinarily with shallow, transverse furrow level with setae ro (missing in M. abeloosi , yet observed in some other Brachypylina ; cf. e.g. Grandjean, 1960a: 348). Postero-lateral regions of inferior wall: with striae bending anteriorly towards plane of symmetry and forming sort of rough, more or less spread reticulation posteromediad; with some longitudinal and transverse ridges prior to reticulated region (not studied in M. abeloosi ).
Dorsovertex. (i.e. posterodorsal region of prodorsum; e.g. Grandjean, 1962b: 79). Differing from M. abeloosi in existence of furrows sf and sd and of ridges ky. Unpaired, curved furrow sf ( Fig. 1 View FIGURE 1 ; intermediary part of sf frequently difficult to see in dorsal projection in LM). Subdividing dorsovertex in two parts: anterior part much smaller and roughly triangular in dorsal projection, and bearing median furrow sm (represented by longitudinal line in Fig. 1 View FIGURE 1 ; also evidenced in M. abeloosi ) and some longitudinal or oblique ridges; according to observations in serial sections, dorsal surface of anterior part of dorsovertex depressed longitudinally and dorsal surface of posterior part appearing anteriorly concave in middle and convex laterally (surface of lamellae slightly convex more abaxially), and posteriorly convex everywhere, with convexity being imperceptibly prolonged by convexity of notogaster (not studied in M. abeloosi ). Oblique furrows sd (not clearly visible in Fig. 2 a). Reached by furrow sf. Both furrows sd rarely meeting together. Ridges ky. Not very frequent and usually bent posteriorly as in Fig. 2 a (not shown in Fig. 1 View FIGURE 1 ).
Lamellae. Sculpture variable. Dorsal surface of lamellae. In M. grootaerti , without striae, yet with generally short and more or less longitudinal ridges often poorly perceptible in LM (example of form and disposition of ridges in Fig. 1 View FIGURE 1 ; with only one relatively long ridge represented on intermediate region of left lamella but, in some other specimens, with two or three long ridges). Dorsal ridges not studied in detail in lectotype of M. abeloosi where, however, some relatively long ridges are present.
Ventral surface of lamellae. With striae and ridges in both species, several elements appearing to prolong striae running onto groove gr. For instance in M. grootaerti , as seen at right in Fig. 1 View FIGURE 1 , among six striae found anteriorly on dorsal wall of gr, two or possibly three running onto anterior, free extension of lamella and, posteriorly, five striae extending to and onto intermediate region of lamella, two striae being distinctly longer. In lectotype of M. abeloosi , with five striae being prolonged on both sides onto anterior extension of lamellae and with possibly nine and ten in intermediate region of lamellae at left and at right, respectively (however, confirmation is required for several prolongations onto inermediate region since no dorso-adaxial limit is distinguishable in lamellae in LM). Note that in both species: (i) ridges running onto posterior part of lamellae are mentioned above; (ii) striae on dorsal and vertical walls of gr arrive at posterior part of upper wall of both carinae tu1 and tu2 of tutoria except some anterior ones (in Fig. 1 View FIGURE 1 , at right, part of anterior stria extending onto lateral region of prodorsum is viewed through lamella and shown as transverse and broken line; in M. abeloosi , striae not running to tu1 are outwardly less numerous but confirmation is required).
Dorso-adaxial limit of lamellae. In M. grootaerti , except posterior portions of furrows sf and sd ( Fig. 1 View FIGURE 1 ; not clearly seen in Fig. 2 a), no precise dorso-adaxial limit is seen in lamellae in usual preparations. Yet, according to observations in serial sections, dorso-adaxial limit seeming to exist in two other places corresponding to bottom of abaxial convexity in anterior region of posterior part of dorsovertex (indicated above) and to furrow (not very apparent in SEM in Fig. 2 a) running ventrally to kx and attenuated posteriorly; note that vertical alignment with internal boundary of lamellae (i.e. spot where dorsal and ventral lamellar walls diverge) is not detected for bottom of furrows sf and sd (internal boundary appears more adaxial) but is observed for furrow ventral to kx at least posteriorly and over short distance.
Tutoria. As in M. abeloosi ( Figs 2 a and 3a of Lions, 1978) and M. senensis ( Fig. 2 a of Bernini, 1975), made up on both sides of three large carinae ( Fig. 2 a and 3a), i.e. two oblique, dorsal carinae tu1 and tu2 located near each other, and carina tu3 more ventrally (note that, owing to position of carinae tu3, leg I probably does not come into contact with tu3 when leg I is folded in groove gr, and therefore carinae tu3 could be more judiciously called ‘subtutorial’).
Carinae tu1. Anteriorly ending in point in both species but, in M. grootaerti , with point smaller and rarely minute or even absent. Thinner and shorter than carinae tu2 (length of tu1 corresponding to 65%, n = 3, of length of tu 2 in M. grootaerti and to 75% on both sides in lectotype of M. abeloosi ). Outline of free border appearing convex over long distance in lateral projection in M.grootaerti , and straight and slightly curved posteriorly in M. abeloosi . Anterior part of tu1: with diversely orientated companion ridges on superior and inferior walls (also evidenced in M. abeloosi ), and with some ridges running to tu2 (anterior, longitudinal ridge extending posteriorly frequently detected); in M. grootaerti , with thin, short line ventrally to point of tu1 (either corresponding to ridge-shaped element or more probably indicating shallow concavity of cuticle observed in SEM just anterior to tutorial elements); in M. abeloosi , with carina running vertically from base of point of tu1 to base of point of tu3 ( Fig. 2 a of Lions, 1978) and, at right in lectotype of M. abeloosi , vertical carina continuous and at left made up of two separate parts (dorsal part seeming to reach bent part of tu2 and to arrive at short ridge arising from longitudinal portion of tu2). With muscle impressions ventrally to middle portion of tu1 (also discernible in M. abeloosi ) and dorsally to posterior portion (not shown in M. abeloosi ). In some instances, tu1 split into two in middle portion; and, in one specimen on one side, with long longitudinal ridge dorsally to tu1.
Carinae tu2. Ressembling M. abeloosi in: (1) anterior ending point lacking in tu2, yet curved ventrally and extending to point of tu3 while thinning; in M. grootaerti and in lectotype of M. abeloosi , at left, ventral extensions touching base of point of tu3 (in lateral projection, free border of extension appearing to be prolonged without gap by dorsal border of point) and, at right in lectotype, extension seeming to finish at abaxial side of point (note that extensions: (i) might come into contact with unguis of legs I when legs I are folded against body; (ii) have intermediate part seldom interrupted ventrally or divided in two); (2) posterior part of tu2 definitely more protruding than in tu1 and tu3; posteriorly, going down sharply and ending at some distance from bothridium (in lateral projection, apparent outline of free border of posterior part of tu2 appearing clearly convex in both species and apparent outline of free border of remaining part ordinarily appearing slightly concave in M. grootaerti and straight on both sides in lectotype of M. abeloosi ); (3) robust and roughly transverse (infrequently oblique) ridges found on superior and inferior walls of tu2 (some ridges appear distinctly bent in lateral projection); (4) existence of some muscle impressions ventrally to intermediate part of tu2. In M. grootaerti , occasionally with one (rarely two) longitudinal, thin and short ridge arising from posterior part of tu2 not far from curve.
Carinae tu3. In both species, always shorter than tu1 and tu2. Anteriorly, usually with ending point (in M. grootaerti and at right in lectotype of M. abeloosi , point generally larger than point of tu1) and, posteriorly, reaching parietal carina kpI (see below and Fig. 2 a). Orientation of tu3: in M. grootaerti , much less oblique than tu1 and tu2; in lectotype of M. abeloosi , as oblique or slightly less oblique. Apparent outline of free border of tu 3 in lateral projection: in M. grootaerti , convex in one or two places, and less often straight or still more seldom concave; in lectotype of M. abeloosi , straight anteriorly and, posteriorly, undulate at left and concave at right. Companion carinae of tu3: in M. grootaerti , frequently two (more infrequently one or three) dorsal and longitudinal ridges and less often one (rarely more) ventral element (occasionally, with some of ridges probably not contiguous to tu3); in lectotype of M. abeloosi : at left, one dorsal ridge and one ventral longer element emerging from middle portion of tu3, bending and running posteriorly and probably not arriving at kpI; at right, one dorso-posterior ridge and two ventral elements longer and curved (postero-ventral companion ridge running to and coming into contact with kpI, and antero-ventral companion ridge divided into two branches, with dorsal branch touching posteroventral companion ridge and with ventral branch reaching kpI). In one specimen on one side, tu3 split into two in middle portion and, in another specimen on one side, tu3 partially weakened.
Bothridia. In both species: opening wide ( Fig. 2 a); unlike dorsal, ventral and posterior parts, anterior part of bothridia not protruding; margin of posterior part of bothridia with well distinguishable notch (larger in M. grootaerti ; not seen in Fig. 1 View FIGURE 1 and partially shown in Fig. 2 a); inside with two curvatures plainly perceptible in apical projection. In M. grootaerti : (1) with auxiliary ridge bent and running parallel to margin of anterior part of bothridia (ridge well visible in lateral projection in Fig. 2 a; in Fig. 1 View FIGURE 1 , shown as very short curved line at right); in M. abeloosi , with thin and oblique line prior to anterior part of bothridia, outwardly forked ventrally at left; (2) according to observations in SEM, on both sides with one to four ventral associated ridges frequently running ventrally to carina ks (see below) and seeming to end near or to arrive at ks; for example, posterior companion ridge extending straight to ks well discernible in Fig. 2 a whereas medial element (also stretching to ks) and anterior element (curved anteriorly and deviating from ks ventrally) being not clearly visible in orientation of Fig. 2 a but better seen in slightly different orientation (ridges have no label in Fig. 2 a); dorsally, one or more ridges sometimes bent anteriorly; not mentioned in M. abeloosi .
Other features. Laterorostral carinae lr. Generally with hiatus observed posteriorly to middle of carinae lr and taking shape of substantial reduction (as found on both sides in lectotype of M. abeloosi ) or possibly of interruption over short distance. Part of lr prior to hiatus: in both species, proceeding from anterior portion of border of camerostome bl.cam and extending close to bl.cam anteriorly and farther from bl.cam posteriorly (due to abaxial curvature of bl.cam; see Fig. 3 View FIGURE 3 a of Lions, 1978, for M. abeloosi ; not shown in M. grootaerti , and anterior segment usually hard to see in ordinary lateral orientation in LM in M. grootaerti ); occasionally with short posteroventral companion ridge (similar, bent element found at left in lectotype of M. abeloosi ).
Part of lr posterior to hiatus: anteriorly, outwardly laminiform and inclined down over some distance (ostensibly over longer distance in M. abeloosi ) and, posteriorly, hidden by ventral part of pedotectum I in lateral and ventral ( Fig. 3 View FIGURE 3 ) projections and not studied in detail; ridges associated to posterior part of lr: (1) in both species, with antero-dorsal ridge emerging either from apparent anterior extremity of posterior part of lr or from short ridge appearing as anterior prolongation of posterior part of lr dorsally to hiatus (as on both sides in lectotype of M. abeloosi ; yet, at right, antero-dorsal ridge possibly not arising from prolongation), running obliquely to and rarely touching tu3, sometimes with one dorsal or ventral associated ridge (antero-dorsal companion ridge not seeming to come into contact with tu3 on both sides in lectotype of M. abeloosi ); (2) in lectotype of M. abeloosi , on both sides, with two short, additional companion ridges running onto acetabular tectum I (cf. Grandjean, 1968: 359–363).
Median bulge. Located in both species on dorsal surface of prodorsum anterior to place where both grooves gr meet, and not very high. In M. grootaerti , mean length = 10.8 µm (range: 9.4–12.0 µm, n = 6) and length in M. abeloosi : 6.4 µm in lectotype and 7.2 µm according to Fig. 3 View FIGURE 3 a of Lions, 1978.
Setae. Rostral setae ro. Mean length = 13.6 µm (range: 12.8–14.9 µm, n = 9) in M. grootaerti and 10.4 µm in M. abeloosi ( Lions, 1978: 406) . Lamellar setae le. Thinning down over overall length instead of only over distal part as in M. abeloosi . Mean length = 21.4 µm (range: 20.4–23.0 µm, n = 6) in M. grootaerti and 15.6 µm in M. abeloosi . Interlamellar setae (in). Situated more posteriorly than in M. abeloosi , and mean length = 10.8 µm (range: 8.5–12.2 µm; n = 8) vs. 5.0–6.0 µm in M. abeloosi . Sensilli bo. Shaped as in M. abeloosi , yet less curved posteriorly (distal bend shown by sensilli variable in M. grootaerti and sensilli often less erect than in lectotype of M. abeloosi ). According to observations in polarized light (not carried out in M. abeloosi ): (1) ending part of sensilli (part inside bothridia not studied) lightening, birefringent, with actinopilin appearing to accumulate in border layer in longitudinal optical section in lateral projection; (2) all examined terminal barbs with proximal portion containing actinopilin and with distal portion isotropic (proximal barbs not studied). Exobothridial setae ex. Thin and smooth, inserted on protuberance (not visible in Fig. 2 a; probably completely lacking at left in lectotype of M. abeloosi ) located at some distance from upper extremity of pedotectum I, and prolonged anteriorly by ridge running obliquely onto inferior wall of pedotectum I and appearing to finish abruptly.
Notogaster ( Fig. 1 View FIGURE 1 ). Apparent outline more convex in lateral projection and more circular in dorsal projection in M. grootaerti . Latero-posterior border tectum absent. According to observations in serial sections in M. grootaerti , notogaster joined to pleural cuticle over some distance posterior to bothridia. Posteriorly to connection between notogaster and pleural cuticle, lateral part of notogaster, which is contiguous to base of pteromorphs, looking first like kind of pad, and more posteriorly changing into lamina appearing narrow, thick (at least, roughly as thick as dorsal tegument of notogaster) and inverted in cross-section (i.e. inclined obliquely towards plane of symmetry). Anywhere else laterally and posteriorly, notogaster with inverted border zone. Note that, in M. abeloosi , inverted border zone is visible posteriorly in dissected notogasters (as shown in Fig. 2 g of Lions, 1978) whereas, in M. grootaerti , inverted border zone is much less distinguishable posteriorly in dissected notogasters as well as in notogaster in not contracted specimens.
Remarkable traits. Lateromedial furrows sl (missing in M. abeloosi , yet detected in M. senensis according to Bernini, 1975: Fig. 2 a). Extending vertically from furrow scd (see below) near latero-abdominal gland gla to well perceptible notch in notogasteral border bng (notch also found laterally on both sides in notogasteral border in M. abeloosi ) and frequently curved anteriorly (in one specimen on one side, furrow sl with lower part substantially bent and ending far from border bng). Remarkable for enlarged borders (enlargement usually appearing better marked dorsally in anterior borders and ventrally in posterior border; in Fig. 1 View FIGURE 1 , furrows sl represented dorsally by anterior border and by line of apparent outline in one place and, ventrally, posterior border looking like very broad convexity).
Lyrifissures. In M. grootaerti : (1) lyrifissures im ( Fig. 1 View FIGURE 1 ): generally appearing in face view as small, often oblong area crossed lengthwise and medially by surface slit (i.e. symmetrical area); frequently darker than surrounding cuticle; note that not very different lyrifissures are observed in another circumdehiscent species, Podacarus auberti Grandjean, 1955 , not closely related to Miracarus (cf. Grandjean, 1955: 112); (2) lyrifissures ia: as usual situated on lower surface of pteromorphs (and accordingly seen through cuticle of pteromorphs in Fig. 1 View FIGURE 1 ); ordinarily similar to roughly rectangular area symmetrical or less often asymmetrical (in asymmetrical areas, surface slit seeming to constitute anterior or posterior limit of area); note that: (i) on both sides in Fig. 1 View FIGURE 1 , surface slit is represented schematically by two parallel broken lines situated side by side, and limit of area is also shown schematically and indicated by broken line; (ii) in orientation of Fig.1 View FIGURE 1 , adaxial part of area is not seen and canal of lyrifissure (not represented in Fig. 1 View FIGURE 1 ) is projected on adaxial part of area); (3) lyrifissures ih and ips: looking like roughly rectangular or triangular area (infrequently irregular in ih) always asymmetrical (surface slit appearing to coincide with ventral limit of area); (4) areas ip of variable form (more frequently irregular or roughly rectangular) and in nearly equal proportions symmetrical or asymmetrical (then surface slit making up posterior limit of area or being close to posterior limit).
In lectotype of M. abeloosi : lyrifissures ia, im and ips at right and ih at left appearing as roughly oblong and symmetrical area; ia, im and ips at left resembling more or less rectangular and symmetrical area; ip at left like rounded and symmetrical area; ih at right and ip at right similar to asymmetrical, more or less oblong area with surface slit being close to ventral and posterior limit of area, respectively. In both species, all areas corresponding to lyrifissures (except possibly areas ia in lectotype of M. abeloosi ) equipped with dots (nature of dots not determined) more frequently observed on limit of areas (note that, in M. grootaerti , dots appear less numerous in areas ia and ip).
Similarities. Elements of humeral regions. Antero-lateral ridges kz (well apparent in Fig. 2 a): associated with carina cir.g; in M. grootaerti , sometimes absent, often shorter than shown at left in Fig. 1 View FIGURE 1 and in Fig. 2 a, and occasionally with auxiliary ridge; in lectotype of M. abeloosi , reaching cir.g comparatively more posteriorly, and less long at left. Lateral furrows sh (represented by line of apparent outline in Fig. 1 View FIGURE 1 ): bearing seta c2 on posterior border in M. grootaerti and in lectotype of M. abeloosi ; with seta c2 closer to cir.g (but probably not in contact with cir.g) on both sides in lectotype.
Pteromorphs (or humeral ailerons; incompletely seen in Fig. 2 a). In both species: looking like narrow lobes in lateral projection and prolonging notogaster without articulation; free border unadorned (e.g. without tooth as in other Microzetidae ; cf. Grandjean, 1936a), yet in M. grootaerti more frequently weakly undulating (more often in posterior part of free border as demonstrated on both sides in Fig. 1 View FIGURE 1 ; also observed at left in lectotype of M. abeloosi ) than crenate. In M. grootaerti : anteriorly, pteromorphs inserted either at side of notogaster or close to free border of lateral notogasteral lamina (see above) whereas, posteriorly, taking root obliquely on surface of notogaster while narrowing (therefore, posteriorly, as shown in Fig. 2 a, lateral notogasteral lamina becoming visible in lateral projection and broadening out progressively; in M. abeloosi , pteromorphs not diverging from border of notogaster posteriorly; compare Fig. 2 a to Fig. 2 g of Lions, 1978); dorsal surface of pteromorphs usually with few ridges (examples given in Fig. 1 View FIGURE 1 ), some of which arriving at cir.g, and ventral surface generally with still fewer ridges or even sometimes without apparent ridge (in lectotype of M. abeloosi , dorsal surface with some ridges orientated longitudinally, and ventral surface with several ridges diversely orientated).
Circumpedal carinae cir.g (according to Grandjean, 1936a: 64, carinae cir.g common in Microzetidae , and posterior legs when folded against body seeming to come partially into contact with notogasteral region ventral to carinae cir.g; capability not verified in M. grootaerti ). Anterior part of cir.g: in both species, appearing clearly curved in lateral projection; in M. grootaerti : according to observations in serial sections, cir.g wholly running onto surface of pteromorphs close to furrow forming dorsal boundary of pteromorphs (furrow not represented in Fig. 1 View FIGURE 1 and not distinctly seen in Fig. 2 a, except outwardly in lower part of ridge kz) and with broadened intermediary portion; and moreover, cir.g anteriorly touching extremity of free border of pteromorphs (in M. abeloosi , location of end of cir.g appearing variable, cir.g reaching anterior extremity of pteromorph on both sides in lectotype but finishing far posteriorly in animal represented by Lions, 1978: Fig. 2 g; in M. senensis , end of cir.g also remote from extremity of pteromorph in animal represented in Fig. 2 a of Bernini, 1975), with intermediary and posterior portions coming closer to free border of pteromorphs than in lectotype of M. abeloosi (less often for anterior portion) and with posterior portion, as seen in Fig. 2 a, frequently fairly parallel to free border of pteromorphs over some distance (also on both sides in lectotype of M. abeloosi ).
Posterior part of cir.g (in Fig. 1 View FIGURE 1 , constituting on both sides apparent lateral border of notogaster): in M. grootaerti , appearing to extend posteriorly in straight line (occasionally slightly undulate) to lateromedial furrow sl in lateral projection, and coming into contact either with ending portion of sl or more often with notch in notogasteral border at which sl arrives (cir.g getting steadily closer to notogasteral border whereas, in M. abeloosi , being parallel to notogasteral border anteriorly and making large curve towards notogasteral border posteriorly); in addition, cir.g terminating more posteriorly than do circumpedal carinae cir.p (see below), except in some instances where, as at left in lectotype of M. abeloosi and as in other Microzetidae ( Grandjean, 1936a: 64) , end of cir.g is approximately on vertical level with posterior extremity of cir.p (at right in lectotype of M. abeloosi , ending portion of cir.g with two associated, probably auxiliary ridges, one running to and seeming to finish near notogasteral notch whereas second associated ridge as well as cir.g ending farther from notch).
Muscle insertions. Antero-dorsal muscle impressions mu.a (corresponding to impressions made by group of muscles which are found with important differences on both sides in anterior region of notogaster in many Circumdehiscentiae; cf. Grandjean, 1966a: 482): in M. grootaerti : frequently demonstrating remarkable arrangement in two oblique rows of three muscle impressions on both sides; positions of muscle impressions not random, yet often appreciably symmetrical in one or several paired elements (rarely none) according to specimens; number of muscle impressions per rows sometimes lower (two elements) or higher (addition of one and more seldom two or three elements); in lectotype of M. abeloosi : four rows of muscle impressions also detected (but only two in specimen in Fig. 1 View FIGURE 1 of Lions, 1978) and made up of three elements (impressions are joined side by side in adaxial row at left), except in abaxial row at right which consists of five muscle impressions; in addition, positions of impressions not symmetrical, except for two pairs of impressions in which symmetry is still poorly pronounced. Latero-dorsal muscle impressions mu.l: located anterior to setae e 2 in both species; in M. grootaerti : of variable size and form; occasionally incompletely delimited; sometimes clearer than surrounding cuticle and, as other muscle impressions, showing more or less large dots (dots are not represented in mu.l as well as in other muscle impressions in Fig. 1 View FIGURE 1 ; nature of dots not determined); in lectotype of M. abeloosi : punctiform at left and looking like reniform, well delimited area without dots at right.
Tendinous insertions. Habitual elements: insertions µ relatively big, with long tendons in both species and located as usual near seta cp; insertions ν with uncommon position in both species (i.e. more or less prior to and abaxially to seta f2 rather than posterior to f2; cf. Grandjean, 1960b: 112), consisting of some marks and without apparent tendons in M. grootaerti , and punctiform and with short tendons in M. abeloosi . Peculiar, lateral insertions (not seen in Fig. 1 View FIGURE 1 ): with long tendons in both species; in M. grootaerti , situated ventrally to muscle impressions of circumdorsal alignment mu.cd (see below) and, in lateral projection, approximately on vertical level with lyrifissure im; in lectotype of M. abeloosi , on both sides, probably in contact with posterior part of muscle impression of circumdorsal alignment mu.cd and located, in lateral projection, roughly on vertical level with seta h3. Circumdorsal furrow scd (not shown in Fig. 1 View FIGURE 1 ). Overlaying muscle impressions of circumdorsal alignment mu.cd as in many other Oribatida ( Grandjean, 1960a: 349) . In M. grootaerti : always shallow and with not clearly defined limits; infrequently completely undiscernible and more often detected laterally on notogaster than posteriorly. In lectotype of M. abeloosi , shallow and perceptible in lateral and posterior regions of notogaster.
Latero-abdominal glands gla (also known as opisthonotal, opisthosomal or oil glands). In both species: relatively small and broad (in M. grootaerti , mean length = 35 µm, range: 29–38 µm, mean breadth = 15 µm, range: 13–18 µm, n = 7; size comparable in M. abeloosi according to Fig. 1 View FIGURE 1 of Lions, 1978); opening of glands on rounded and slightly protruding tubercle (occasionally not distinguishable in M. grootaerti ) In M. grootaerti , with small and generally distinctly delimited convexity in internal surface of cuticle ventrally to opening of gland gla.
Dissimilarities. Line of large scissure. In M. grootaerti , suppressed superficially in median region of notogaster (in contrast, well distinct everywhere in M. abeloosi ); yet, in median region of notogaster, with two convex lines observed in depth in dorsal projection: anterior line ordinarily discernible only at high magnification (accordingly not seen in Fig. 1 View FIGURE 1 ), and posterior line usually more or less visible at low magnification (whether parting between prodorsum and notogaster corresponds to anterior line or posterior line is unclear). Lateral, perceptible parts of line of large scissure sometimes shorter and rarely longer than at left and at right, respectively, in Fig. 1 View FIGURE 1 . Notogaster strongly joined to prodorsum in M. grootaerti : even after heating for ca. 20–30 minutes in lactic acid, notogaster always hard to separate from prodorsum in dissections and moreover separation invariably resulting in break of cuticle; for example, after separation of notogaster from prodorsum in two specimens, tear is found in cuticle of prodorsum in one specimen, and in cuticle of prodorsum on one side and of both prodorsum and notogaster on other side in second specimen; in addition, splitting of cuticle into two layers is detected in one place on one side in both specimens (study not made in M. abeloosi ).
Muscle impressions of circumdorsal alignment mu.cd. Remarkable in M. abeloosi because of lateral boundaries appearing to be prolonged ventrally, suggesting that impressions mu.cd are passably broader than in general (yet confirmation is required). Prolongations more distinguishable when seen obliquely, and appearing shorter in muscle impressions located anteriorly; in muscle impressions of dorso-posterior group prolongations outwardly missing.
Other characters: (1) anterior carinae kw (in Fig. 1 View FIGURE 1 , represented only at right; undiscernible in Fig. 2 a) infrequently found close to and more or less parallel to line of large scissure (absent in M. abeloosi ); (2) location of lyrifissures ih and ips: in M. grootaerti , ih situated on posterior border of furrows sl, and ips borne on vertically developed bulge (in dorsal projection, apparent lateral outline of bulge either accentuated, as at left in Fig. 1 View FIGURE 1 , or less frequently not very marked or indistinguishable as shown at right in Fig. 1 View FIGURE 1 ; bulge not detected in M. abeloosi ); distance between lyrifissures ih and ips variable in both species: in M. grootaerti , mean distance in lateral projection = 15.5 µm (range: 9.5–24.0 µm, n = 10); in M. abeloosi , 7.0 µm in specimen in Fig. 2 g of Lions (1978) and ca. 16.0 µm on both sides in lectotype, respectively; (3) racemiform organs not seen in M. grootaerti ; (4) fecundity: no gravid females collected (n = 7) with more than two eggs as in other Microzetidae ( Grandjean, 1936a: 65) .
Setae. As in other Microzetidae ( Grandjean, 1936a; Travé, 1956; Lions, 1966), notogasteral setae consisting of 9 pairs, i.e 10 pairs corresponding to ordinary number in Brachypylina (cf. Grandjean, 1959a: 178) minus pair of setae p3, and anterior pair of series c regarded as pair of setae c2 owing to location in humeral region. Yet, contrary to some Microzetidae (e.g. Nellacarus petrocoriensis Grandjean, 1936 ), setae e2 more distant from plane of symmetry than setae f2 (however, in M. grootaerti , distance between e2 and plane of symmetry never as short as distance at right and occasionally longer than distance at left in M. abeloosi specimen represented in Fig. of Lions, 1978). In M. grootaerti , except pseudanal setae p1 which are erect, notogasteral setae are bent small, thin and smooth as in M. abeloosi ; and, unlike M. abeloosi , setae h3 are never clearly anterior to opening of glands gla (according to Grandjean, 1961b: 307, position of h3 appearing unusual in M. abeloosi as also in several other Microzetiade, as e.g. Phylacozetes membranulifer Grandjean, 1936 ). Note that Dometorina notations modified by Lions (1970) are used here for labelling notogasteral setae.
Lateral characters ( Fig. 2 a). In both species, pleural regions of podosoma without distinct sejugal furrow and without custodium. With deep sejugal depression posterior to bothridia; in M. grootaerti , according to observations in serial sections, sejugal depressions: (1) developed in pleural regions dorsal to legs, with dorsal boundary formed to great extent by carinae kl (see below), and finishing posteriorly level with intermediate part of carinae kl; (2) over most of length, apparent transverse outline: broadly and more or less V-shaped; dorsally straight and making obtuse angle with plane of symmetry; ventrally either convex or straight (then appearing roughly parallel to plane of symmetry); showing furrow-like bottom; (3) accompanied by carinae kb (see below) running over long distance dorsally to furrow at bottom of sejugal depressions (accordingly, in Fig. 2 a, furrow is perceptible just ventral to base of carina kb). Study of following lateral characters completes description of lateral elements of prodorsum.
Remarkable traits. Dorsal carinae kl. Long and oblique, and deviating perceptibly posteriorly from free border of ventral shield. In both species, made up of three parts. Anterior part of kl: precise, with distinct crest line, and extending to pteromorphs (according to observations in serial sections in M. grootaerti , extremity of kl appearing situated on or near base of pteromorphs posteriorly to bothridium). Intermediate part of kl: broadened, prominent and convex in dorsal projection (part relatively less apparent in dorsal projection in lectotype of M. abeloosi ): (1) in M. grootaerti : with one to three hollows (two hollows labelled 2 in Fig. 2 a) in dorsal wall, of variable form and with well pronounced margin usually appearing to be partially interrupted; with some muscle impressions detected dorsally to and ventrally to hollows (according to observations in serial sections, muscles probably also fixed level with hollows); (2) in M. abeloosi according to Lions (1978: 411 and Figs 2 h, 2i and 3a): with slit ‘protected’ by two curved ridges (one ventral and one dorsal) and with muscle impressions found ventrally to lower ridge (in lectotype, slit might be absent or blocked since only roughly oval outline is detected on both sides between crest line of kl and dorsal associated ridge; on both sides, both associated ridges appearing to reach crest line of kl, and ventral associated ridge straight at right and shorter at left than in Fig. 2 i of Lions, 1978). Posterior part of kl: running to and arriving at some distance from extremity of cir.p: (1) in M. grootaerti : mean distance between junction cir.p-kl and extremity of cir.p = 16.4 µm (range: 7.7–20.8 µm; n = 6); anteriorly, often with longitudinal associated ridge (for example, in Fig. 2 a, ridge found dorsoposteriorly to second hollow and, as frequently happens, not seeming to come into contact with hollow); posteriorly, in lateral projection ( Fig. 2 a), crest line growing feebler swiftly and becoming suddenly indistinct; (2) in lectotype of M. abeloosi , crest line completely but poorly distinguishable at right and recognizable only near cir.p at left in lateral projection; furthermore, kl touching closer extremity of cir.p at left (distance between junction cir.p-kl and extremity of cir.p = 14.2 µm and 21.8 µm at left and at right, respectively).
Subdorsal, oblique carinae kb (missing in M. abeloosi ). Consisting of two parts separated either by only one curve (which can be more clearly defined than in Fig. 2 a) or more frequently by two more or less marked curves; note that portion of carinae kb between both curves: (i) averages 12.2 µm long (range: 8.7–14.5 µm, n = 6) in lateral projection; (ii) is frequently projected on base of pleurophragma hl (see below) in lateral orientation (less often only one of separating curves or single separating curve is projected on base of hl); (iii) is represented schematically, on both sides in Fig. 1 View FIGURE 1 , as weakly bent broken line because carinae kb are very poorly detectable through cuticle of notogaster in dorsal view level with pleurophragma hl. Anterior part of kb: rarely absent; running anteriorly close to carina kv (see below), spacing between kb and kv being often narrow as in Fig. 2 a; ending near and dorsally to extremity of kv or more frequently reaching kv (not clearly shown in Fig. 2 a); with two associated companion carinae originating either from or just posterior to single separating curve or, when two separating curves are present, from portion of kb between both curves, and consisting of dorsal ridge (hardly discernible in Fig. 2 a) extending obliquely to dorsal part of bothridium (ending portion of dorsal ridge hidden by pteromorph in lateral projection and not studied) and of occasional, short and ventral ridge (not seen in Fig. 2 a; in few instances, origin of dorsal ridge seeming to coincide with origin of ventral ridge). Posterior part of kb: finishing abruptly at some distance from carina kh (see below); sometimes with associated ridges (not shown in Fig. 2 a) as e.g. dorsal and more or less parallel, auxiliary element; note that: (i) posterior part of kb appears to be made up, in one specimen on one side, of three separate elements disposed one after another, and in another specimen on one side, of two separate elements, posterior element running dorsally to anterior one over some distance; (ii) V-shaped ridge visible in Fig. 2 a ventrally to ending portion of kb appears uncommon.
Parietal carinae kpII and careniform ribs on acetabular tecta II (carinae kpII probably not very frequent in Brachypylina according to survey of data in Grandjean’s works and in works of some other authors; yet see Grandjean, 1960c: 263, for example in another brachypylinid species; outwardly present in M. senensis according to Fig. 2 a of Bernini, 1975). Dorsal part of kpII (missing in lectotype of M. abeloosi ): in M. grootaerti : sometimes absent; according to observations in SEM, ending portion of kpII arriving posteriorly at dorsal end of free border of acetabular tectum II (i.e. dorsal commissure j.d. of acetabular tectum sensu Grandjean, 1968: 360), and common extremity located either dorsally to or more often posteriorly to trochanter II; note that: (i) when common extremity is posterior to trochanter II as shown in Fig. 2 a, ending portions of kpII and of free border of acetabular tectum are longer and distinctly bent ventrally, and common extremity is generally hidden in lateral projection by carina 4 associated with pedotectum II (see below); (ii) in lectotype of M. abeloosi , ending portion of free border of acetabular tectum II is longer and curved over longer distance at left and, on both sides, commissure j.d. is visible in lateral projection; (iii) in M. grootaerti , opening z of ductus of coxal glands (frequently situated dorsally to legs II in oribatid mites) is observed between crest line of kpII and free border of acetabular tectum II ( Fig. 2 a); with comparable location in M. abeloosi just dorsally to free border of acetabular tectum II (see Fig. 3 View FIGURE 3 a of Lions, 1978).
Intermediate part of kpII ( Figs 2 a and 3a): in M. grootaerti : in contact with some acetabular ribs; often with one (more seldom two) associated carina ventral to lower acetabular rib and either oblique (e.g. slightly sinuous companion ridge represented in Fig. 3 View FIGURE 3 a; in one specimen on one side, with longer companion ridge bent at angle; in another specimen on one side, with two parallel auxiliary ridges, dorsal one being distinctly shorter) or transverse (and then finishing near cir.p); in lectotype of M. abeloosi : outwardly touching all ribs at right and several ribs at left; dorsally, on both sides, making anterior half loop (larger at left); ventrally, at left, curved and extending posteriorly over short distance before finishing abruptly and, at right, reaching posterior portion of lower acetabular rib; on both sides with associated, mainly auxiliary carinae probably coming into contact with cir.p, namely: (1) at left, four ridges: anterior, oblique and S-shaped ridge ventral to lower acetabular rib and partially divided into two; other ridges transverse and ventral to anterior ridge except posterior ridge which extends to and arrives at dorsal extremity of anterior ridge before touching ending portion of kpII posteriorly to ventral curve (posterior ridge partially split into two); (2) at right, five ridges transversally orientated: both anterior and medial ridges seeming to end dorsally close to lower acetabular rib (medial ridge and ridge posterior to medial one nearly completely divided into two).
Ventral part of kpII (indistinguishable in lectotype of M. abeloosi ): in M. grootaerti : frequently present (but missing in Fig. 3 View FIGURE 3 a) and occasionally well developed (and then more often running to cir.p than to pedotectum II, outwardly without reaching any one); sometimes with one or two, anterior or posterior auxiliary ridges frequently extending transversally (note that, on one side in two specimens in which ventral part of kpII is lacking, short longitudinal ridge is detected between ending portion of kp II and free border of acetabular tectum II).
Acetabular careniform ribs (3 Fig. 2 a): in M. grootaerti : consisting of six to 14 often robust elements; one or two upper ribs appearing to be ordinarily located dorsally to acetabulum II in lateral projection, and some others extending anteriorly onto superior wall of pedotecta I; in M. abeloosi : number and size of ribs variable ( Lions, 1978: 411): for example, in lectotype, with eight ribs at right (three relatively long; none appearing dorsal to acetabulum II in lateral projection; outwardly only one running onto superior wall of pedotectum I) and nine ribs at left (five relatively long; two appearing dorsal to acetabulum II in lateral projection; three extending onto superior wall of pedotectum I); in addition, posterior portion of lower acetabular rib, at left, arriving at ventral curve of kpII and, at right, running to and finishing at short distance from free border of acetabular tectum II.
Crooked carinae kg. Dorsal part of kg: shorter than carinae kf (see below) in M. grootaerti and in lectotype of M. abeloosi ; in M. grootaerti : either transverse or more frequently oblique and more or less parallel to kf ( Fig. 2 a); absent in one specimen on one side; in lectotype of M. abeloosi , diverging from kf on both sides. Ventral part of kg: in M. grootaerti : making markedly rounded curve on dorsal region of acetabular tectum III (note that, in both species, dorsal region of acetabular tectum III is broadened, with free border distinctly curved) and, according to observations in SEM, coming into contact with free border of acetabular tectum; in lectotype of M. abeloosi : on both sides, straight and forming large obtuse angle with dorsal part of kg, and probably not touching free border of acetabular tectum III. Carinae associated to kg in M. grootaerti (all located posterior to kg) consisting of (frequencies not estimated): (1) one or two companion ridges extending dorsally, forked or not (not seen in Fig. 2 a; not observed in lectotype of M. abeloosi ); (2) one vertical or rarely oblique companion ridge (7 in Fig. 2 a) in contact with kg over more or less long distance and reaching extremity of crest line of protrusion R (see below); note that: (i) in two specimens on one side, element 7 is not connected with protrusion R, and is L-shaped in one specimen and U-shaped with branches directed ventrally in second specimen; (ii) in lectotype of M. abeloosi , at right, ridge 7 deviating from kg both dorsally and ventrally, and outwardly absent at left; (3) one (more infrequently two) longitudinal and more or less bent, companion or auxiliary ridge situated dorsally to crest line of ending portion of R (element hardly visible in Fig. 2 a, yet better perceptible in slightly different orientation; not detected in lectotype of M. abeloosi ). Note that both ridges represented on acetabular tectum III in M. abeloosi in Fig. 3 View FIGURE 3 a of Lions (1978) are not found in lectotype.
Acetabular protrusions R and associated carinae. Protrusions R: conspicuous, cariniform elements on acetabular tectum III; apparent outline appearing angular with rounded top in ventral projection ( Fig. 3 View FIGURE 3 a); note that, according to observations in serial sections: (i) well marked bulge created by acetabular tecta III forms dorsolaterally prominent carina which appears to carry protrusion R anteriorly and to prolong R posteriorly; (ii) cuticle of pleural region exhibits gutter-like depression (ca. 20 µm high level with middle region of R) just dorsally to acetabular bulge; (iii) posteriorly, dorsolateral acetabular carina accompanied by pleural depression extends beyond acetabulum and to discidium (see below). Distance measured in ventral projection ( Fig. 3 View FIGURE 3 a) between top of protrusions R and top of discidia: mean value = 16.6 µm in M. grootaerti (range: 14.3–19.8 µm, n = 10); in M. abeloosi , distance of ca. 11.0 µm according to Fig. 2 c of Lions, 1978, and of 13.7 µm and 14.3 µm in lectotype; note that both tops are approximately equidistant from plane of symmetry in both species. Crest line of R: in M. grootaerti ( Fig. 2 a), arriving at carina kg or less frequently either ending more or less close to kg or, as mentioned above, coming into contact with ridge 7 associated with kg; in lectotype of M. abeloosi , finishing at short distance from carina kg at left and distinctly farther from kg at right. Crest line of extension of acetabular carina posterior to R: either arriving at superior wall of discidium (yet possibly sometimes not reaching discidium; unclear in Fig. 2 a but, according to slightly different orientation, crest line appearing to come into contact with cuticular granule located dorsally to carina 9 mentioned below) or less often touching carina kd associated with discidium (see below); in lectotype of M. abeloosi , probably arriving at discidium at left and ending close to kd at right.
Regular, dorsal associated carinae: (1) in M. grootaerti : consisting of anterior carina 8 usually situated level with top of R and of posterior neighbouring carina (both elements finishing close to or at crest line of R; in one specimen on one side, posterior carina clearly curved posteriorly and possibly reaching crest line of extension of acetabular carina posterior to R); anterior carina 8: with ventral part generally appearing broadened in lateral projection; ordinarily relatively long and then with dorsal part frequently divided into two branches (rarely three or four) extending to and occasionally arriving at kh; posterior carina: either short or long and then curved or more rarely sinuous or straight and divided into two branches (as possibly in Fig. 2 a) or not; with ventral part often broadened in lateral projection; when posterior carina (or branches of posterior carina) is well developed, running either to kh or to kd or to dorsal commissure of acetabular tectum IV or to widening o.d. of acetabular tectum (i.e. space located ventrally to acetabular tectum between border of circumtrochanteral opening and dorsal commissure j.d. of tectum; cf. Grandjean, 1968: 362); (2) in lectotype of M. abeloosi : element equivalent to anterior carina in M. grootaerti at right with two oblique branches extending to and terminating near kh and near dorsal commissure of acetabular tectum IV, respectively; shorter and oblique at left; element equivalent to posterior carina in M. grootaerti with two branches at right (anterior oblique branch running to and ending near widening o.d. of acetabular tectum IV; posterior longitudinal branch extending to and probably coming into contact with kd) and with three oblique branches at left (dorsal branch going to and touching kh; medial branch straight, running to and finishing near kh; ventral branch extending to and reaching dorsal commissure of acetabular tectum IV); broadening of ventral part observed in anterior and posterior elements in lateral projection.
Less frequent, additional dorsal associated carinae: (1) prior to carina 8: in M. grootaerti , one or less often two ridges (or even three in one specimen on one side) frequently arriving at crest line of R; either short and vertical or longer and bent; running to and usually not touching kh (in Fig. 2 a, two additional ridges might be present, anterior one appearing short and straight, and posterior one longer and curved); in lectotype of M. abeloosi , at right, one short and broad ridge in contact with crest line of R (absent at left); (2) posterior additional carinae: in M. grootaerti , made up of ridge of variable form, orientation and size (not studied in detail, yet more often roughly straight and orientated anteriorly and obliquely; not clearly perceptible in Fig. 2 a) and, more posteriorly, of ridge short, generally curved and reaching kd (in Fig. 2 a, posterodorsal additional carina not very discernible, but nonetheless appearing to arise from crest line of extension of acetabular carina posterior to R); in lectotype of M. abeloosi , posterodorsal additional ridges not arriving at crest line of R and consisting of one element at right (short and more or less transverse) and of two elements at left (anterior ridge longitudinal, extending to and outwardly ending close to auxiliary ridge of kd mentioned below; posterior ridge longer, directed obliquely and parallel to crest line of R).
Ventral carinae kt (poorly visible in Fig. 2 a; dorsal ending part generally not very perceptible in LM in lateral projection): (1) in M. grootaerti : emerging from top of R (or, if not, probably slightly posteriorly); broadly bent anteriorly (curvature appearing regular except in some instances where kt is either sinuous or outwardly split into two over limited distance); running to and frequently coming into contact with cir.p ( Fig. 3 View FIGURE 3 a); ordinarily with one roughly parallel associated ridge of variable size, located anteriorly to or more often posteriorly to kt, sometimes originating from kt, and touching cir.p or not (difficult to see in Fig. 2 a; in Fig. 3 View FIGURE 3 a, short ventral auxiliary ridge reaching cir.p discernible posteriorly to kt); in one specimen on one side, with two posterior associated ridges: dorsal ridge proceeding from kt, and ventral ridge arriving at cir.p; (2) in lectotype of M. abeloosi , on both sides: kt broadly curved and extending from top of R to cir.p; with anterior, associated ridge (at right, associated ridge touching dorsal part and probably also ventral part of kt; at left, associated ridge might be auxiliary element running from crest line of R to cir.p and being ventrally very close to kt).
Less frequent, additional ventral associated carinae: (1) anterior to kt: in M. grootaerti , short and oblique ridge usually arising from crest line of R, and bent or not (in SEM in Fig. 2 a, appearing more luminous than adjacent cuticle, and line seen ventrally corresponding either to line of apparent outline or to associated ridge directed posteriorly; in one specimen on one side, ridge clearly longer; in Fig. 3 View FIGURE 3 a, ridge not distinguishable); no equivalent element detected in lectotype of M. abeloosi ; (2) posterior to kt: in M. grootaerti , one to three ridges of variable size (for example, two ridges are shown in Fig. 2 a and three in Fig. 3 View FIGURE 3 a; in Fig. 2 a, anterior element directed obliquely and posteriorly, and seeming to finish abruptly at some distance from posterior ridge 9 which is located near extremity of extension of acetabular carina posterior to R; in Fig. 3 View FIGURE 3 a, posterior element also labelled 9 and, possibly like medial element, appearing to emerge from extension of acetabular carina posterior to R); posteroventral additional ridges usually not coming into contact with cir.p; in lectotype of M. abeloosi , at left, posteroventral additional ridge appearing roughly parallel to crest line of R, and absent at right.
Carinae and tecta. Pedotecta I and associated carinae. Pedotecta I ( Figs 1 View FIGURE 1 , 2 a and 3a): in both species, large, with distal limbus, with free border extending posteriorly beyond exobothridial seta ex, and with base (i.e. proximal limit of inferior wall of pedotecta) showing broad, anterior convexity visible through cuticle in lateral projection. Carinae on superior wall of pedotecta I (ridges borne by intermediary region of superior wall of pedotecta I as well as by inferior wall of pedotecta I not studied): (1) carinae ku ( Figs 1 View FIGURE 1 , 2 a and 3a): in both species, large and clearly curved; (2) ridges found in dorsal region of pedotecta I: fewer in M. grootaerti (note that long ridge parallel to free border of pedotectum, as seen in Fig. 2 a, is infrequent but not exceptional); in lectotype of M. abeloosi , made up of seven elements at left and of six elements at right (note that more elements are represented in Fig. 3 View FIGURE 3 a of Lions, 1978); (3) J-shaped carinae cl ( Fig. 3 View FIGURE 3 a): anterior part of cl running onto antero-ventral region of podosoma: in M. grootaerti , either ending near or more often reaching abaxial branch of V-shaped carina cv of mentotectum (see below), and junction between cl and branch of cv frequently appearing rounded and attenuated in ventral projection; in lectotype of M. abeloosi , at right, arriving at ridge extending obliquely onto mentotectum and, at left, finishing close to transverse part of podocephalic fissure F (see below), without contact with oblique ridge of mentotectum; posterior part of cl running onto pedotectum I: outwardly terminating abruptly on both sides in M. grootaerti and at left in lectotype of M. abeloosi whereas, at right in lectotype, appearing longer and touching ventral ending portion of ridge extending obliquely onto intermediate region of pedotectum; ridges associated to cl consisting: in M. grootaerti , of one or less frequently two or three usually companion elements generally bent (one anterior companion ridge is seen in Fig. 3 View FIGURE 3 a); in lectotype of M. abeloosi , of one anterior ridge at right and one posterior ridge at left, both ridges arriving at cl; note that: (i) in M. grootaerti , cl is rarely reached by anterior prolongation of cir.p which is occasionally observed; (ii) in lectotype of M. abeloosi , cir.p extends anteriorly on both sides but only prolongation at left appears to touch cl.
Carinae in contact with dorsal ending part of free border of pedotecta I ( Fig. 2 a; lacking in M. abeloosi ): (1) carinae ks: short and ordinarily curved and accentuated; going to and either ending near or infrequently seeming to reach kv; (2) carinae kk: straight or bent (infrequently bent at angle); rarely poorly perceptible in lateral projection in LM; running obliquely and posteriorly either to acetabular ribs (and then probably sometimes touching uppermost rib; in one specimen on one side, kk coming into contact with rib located ventrally to uppermost rib whereas, in another specimen on one side, kk arriving at posterodorsal prolongation of still more ventral rib) or less often to kv (and then finishing at some distance from kv). Ventral ridges posterior to carinae ku ( Fig. 3 View FIGURE 3 a): in M. grootaerti , made up of one or two ridges (seldom three, and rarely none) of variable size (long elements appearing to be infrequent) and orientation (for example, Fig. 3 View FIGURE 3 a shows three ridges: one transverse element near carina ku and, more posteriorly, one oblique and one longitudinal element); note that: (i) overall, oblique ridges are more frequent; (ii) occasionally longitudinal ridges might arrive at pedotectum II whereas oblique and transverse ridges appear to come into contact with cir.p or junction ku-cir.p; in lectotype of M. abeloosi , only ridges associatd to kpII as indicated above are found posterior to ku.
Median carinae kv (not shown in Fig.1 View FIGURE 1 ). In M. grootaerti : dorsal portion of kv curved anteriorly; extending to bothridium and, after having reached kb (as mentioned above) or not, ending either posteriorly to bothridium or more seldom at posterior part of bothridium; ventral portion of kv slightly bent (and then kv appearing S-shaped in lateral projection as seen in Fig. 2 a) or not; running to and finishing at some distance from pedotectum II (sometimes not far from carina 4 associated with pedotectum II; in one specimen on one side, probably touching parietal carina kp II); intermediary portion of kv infrequently reduced or outwardly absent. Main differences in lectotype of M. abeloosi : less robust; at left, dorsal portion bent at an angle, with dorsal and oblique auxiliary ridge and possibly also with short, longitudinal companion element going to ventral part of auxiliary ridge; on both sides, ventral portion extending transversally to trochanters II and terminating far from free border of acetabular tecta II.
Parietal carinae kpI (common in Brachypylina according to Grandjean, 1959b: 458). Part of kpI dorsal to carinae tu3: oblique and curved posteriorly; in M. grootaerti and at right in lectotype of M. abeloosi , ending abruptly at some distance from base of pedotectum I (ending portion of kpI occasionally bent dorsally in M. grootaerti and inclined ventrally at right in lectotype) whereas, at left in lectotype, appearing to be prolonged posteriorly (see below). Carinae associated to dorsal part of kpI: (1) in M. grootaerti : with robust longitudinal companion carina (rarely absent or reduced) just dorsal to carina tu3 ( Fig. 2 a), bent ventrally or not, and either not arriving at tu3 (and then growing feebler anteriorly over more or less long distance before finishing) or coming into contact with posterior part of tu3 (and then, with tu3 and with kpI, delimiting conspicuous, complete or incomplete concavity of variable form); less frequently with another longitudinal companion carina dorsal to previous companion element, either separated from previous companion element or, with previous companion element and with kpI, delimiting second concavity; (2) in lectotype of M. abeloosi , with two ridges at left and outwardly with four ridges at right originating dorsally from kpI and running transversally over short distance onto adjacent cuticle; (3) in both species, with posterior and oblique ridge reaching convexity of base of pedotectum I: in M. grootaerti , either arriving at extremity of kpI (ending portions of both elements making then broad angle) or less often finishing close to extremity of kpI or at underside of ending portion of kpI or far from kpI; in lectotype of M. abeloosi , touching underside of kpI at right and appearing as posterior prolongation of kpI at left; note that: (i) in M. grootaerti , posterior ridge frequently goes over short distance up inferior wall of pedotectum I and, on both sides in lectotype of M. abeloosi , posterior ridge probably comes into contact with long carina extending vertically onto inferior wall of pedotectum I; (ii) in both species, dorsal commissures j.d. of acetabular tecta I are located on convexity of base of pedotecta I just ventrally to posterior ridge.
Part of kpI ventral to carinae tu3: (1) in M. grootaerti : dorsally, generally directed obliquely and posteriorly ( Fig. 2 a) and, more ventrally, either running straigth to free border of acetabular tectum I (kpI might reach free border of acetabular tectum, yet extremity of kpI is always difficult to see) or curved anteriorly and then either arriving at part of laterorostral carina lr posterior to hiatus (see above) as shown in Fig. 3 View FIGURE 3 a or more often terminating more or less close to lr (then ending portion of kpI sometimes forming posterior border of concavity which is delimited in other places, completely or incompletely, by anterior companion ridge associated with ending portion of kpI); (2) in lectotype of M. abeloosi : on both sides, ventral part of kpI going to dorsal additional ridge associated with lr posterior to hiatus (see above); at left, coming into contact with dorsal ridge associated with lr; at right, bending anteriorly at some distance from dorsal ridge associated with lr and touching lr.
Pedotecta II and associated carinae. Pedotecta II relatively large and forming complete limbus. Dorsal, lowered part of pedotecta II: extending to carina kf; in M. grootaerti : bent or roughly straight (and then with ending portion curved or not) or less frequently slightly sinuous (in Fig. 2 a, with dorsal portion straight and with ventral portion outwardly bent); reaching or more often ending near kf (then mean distance between extremity of dorsal part of pedotecta II and kf in lateral projection = 2.8 µm, range: 2.2–4.5 µm, n = 10); in lectotype of M. abeloosi , curved at right and slightly sinuous at left, and distance between extremity of dorsal part of pedotecta II and kf = 5.5 µm at right and 2.5 µm at left. Carinae associated with inferior wall of pedotecta II consisting of: (1) ventral companion ridges (in M. grootaerti , one or two, two being represented by broken line in Fig. 3 View FIGURE 3 a; two at right and three at left in lectotype of M. abeloosi ); (2) some other ridges (not studied in detail; one is labelled 4 in Fig. 2 a; in M. grootaerti , carinae 4 either parallel to or drawing near to dorsal part of pedotecta II; in lectotype of M. abeloosi , ridge equivalent to carina 4 in M. grootaerti deviating from dorsal part of pedotectum II on both sides).
Dorsal carinae associated with superior wall of pedotecta II made up of: (1) in M. grootaerti , one, two or three companion or auxiliary ridges straight or more frequently bent posteriorly (two elements, labelled 5, are seen in Fig. 2 a; when three elements are present, following destinations reached or not are observed: for dorsal ridge, kf or more seldom antero-ventral parallel ridge associated with kf as mentioned below; for medial ridge, kf or more often dorsal commissure j.d. of acetabular tectum III; for ventral ridge, commissure j.d. or more frequently adaxial wall of dorsal widening o.d. of acetabular tectum (i.e. ‘floor’ of widening according to Grandjean, 1968: 362); (2) in lectotype of M. abeloosi : four ridges at left including three dorsal elements (two uppermost elements running to and probably arriving at kf while other element going to but not touching dorsal commissure of acetabular tectum III) and one ventral and auxiliary element less oblique and extending to and finishing at some distance from floor of dorsal widening of acetabular tectum III; one ridge at right (dorsally running to and coming into contact with dorsal commissure of acetabular tectum III and ventrally forked).
Ventral carinae associated with superior wall of pedotecta II comprising ridge 6, carina kq and ridge in vicinity of coxisternal seta 3c. Ridge 6: in M. grootaerti (poorly distinguishable and partly hidden in Fig. 2 a; not shown in Fig 3 View FIGURE 3 a): infrequently absent; not long (maximum length of ca. 13.0 µm measured in one specimen on one side); oblique (rarely longitudinal) and straight or curved (seldom weakly sinuous); anteriorly, ending abruptly at some distance from free border of pedotectum II (extremity of ridge 6 generally farther from free border of pedotectum II than extremity of carina kq mentioned below), and infrequently reaching either kq or dorsal associated ridge of kq; posteriorly, going to anterior tooth Δ ’ of podosoma-trochanter articulation III and either finishing near Δ ’ or less often extending to and arriving at floor of ventral widening o.v. of acetabular tectum III; sometimes with dorsal associated ridge (frequently companion ridge proceeding from intermediary part of 6) finishing abruptly at some distance from 6; in lectotype of M. abeloosi : anterior part of 6 forked at left; on both sides, 6 running posteriorly first to anterior articular tooth Δ ’ III and then deviating towards and touching floor of ventral widening of acetabular tectum III, and with anteroventral companion element extending also posteriorly to floor of ventral widening of acetabular tectum III (at right, arriving at floor of ventral widening and seeming to come into contact with posterior extremity of 6 and, at left, growing feebler and probably not reaching floor of ventral widening).
Carina kq: in both species, more ventral and longer than 6, and posteriorly arriving at floor of ventral widening of acetabular tectum III (with few exceptions in M. grootaerti ); in M. grootaerti (not shown in Fig. 2 a; note that, in orientation of Fig. 3 View FIGURE 3 a, dorsal free border of pedotectum II is partially hidden by kq and hidden portion is partly represented by short broken line anterior to kq while another portion of dorsal free border of pedotectum II seen by transparency through trochanter III is also indicated by broken line): kq oblique and either bent or less frequently slightly sinuous (rarely straight); with anterior extremity ordinarily projected in lateral orientation approximately on base of pedotectum II; with associated ridges involving one (occasionally two or seldom three) dorsal and more often auxiliary element (usually oblique and rarely vertical; more frequently straight; often shorter than kq; sometimes touching floor of ventral widening of acetabular tectum III) and occasionally one (rarely two) ventral and companion or auxiliary element (curved or straight; of variable orientation; shorter than kq; note that ridge arising from pedotectum II and shown in Fig. 2 a dorsal to coxisternal seta 3c might be infrequent, ventral element associated with kq).
In lectotype of M. abeloosi , on both sides, kq better marked than in M. grootaerti and slightly sinuous; with anterior extremity being projected in lateral orientation anteriorly to base of pedotectum II; with associated companion ridges: composed, on both sides, of anterodorsal ridge (running anteriorly while diverging moderately from kq; simple at right; divided into two branches at left: dorsal branch appearing to be approximately as long as right anterodorsal ridge; ventral branch longer and possibly coming into contact with free border of pedotectum II) and of posteroventral ridge (bent posteriorly and with curvature farther from kq at right; extending to and reaching floor of ventral widening of acetabular tectum III, at right, and running to and arriving at ventral commissure j.v. of acetabular tectum III, at left); and moreover consisting, at right, of ventral ridge (comprising short upper portion, and oblique and longer lower portion extending vertically to and finishing near cir.p) and, at left, of posterodorsal ridge (bent posteriorly; running to and touching floor of ventral widening of acetabular tectum III).
Ridge in vicinity of coxisternal seta 3c: in M. grootaerti : infrequently lacking; ordinarily adaxial to seta 3c ( Fig. 3 View FIGURE 3 a); mean length = 8.0 µm in ventral projection (range: 5.6–10.3 µm, n = 7); most often longitudinal and straight; sometimes appearing to come into contact with cir.p anteriorly; in lectotype of M. abeloosi , ridge: at left, long and roughly U-shaped, with branches directed anteriorly and outwardly extending from free border of pedotectum II dorsally to cir.p ventrally (with two setae 3c, posterior one appearing to be borne by ridge); at right, shorter and V-shaped, with branches directed dorsally and finishing abruptly approximately on longitudinal level with 3c which is located between both branches.
Discidia and associated carinae. Discidia (top labelled dis in Figs 2 a and 3a): laminiform, triangular and oblique elements very prominent in both species. Anterior part of discidia: (1) in M. grootaerti : with free border usually running obliquely and steadily to plane of symmetry and reaching cir.p ( Fig. 3 View FIGURE 3 a); note that free border appears to be bent in ventral and apical view (and then free border apparently consisting of two portions, slope of adaxial portion being distinctly more pronounced relative to plane of symmetry), in one specimen on one side, and in another specimen on one side, ends abruptly dorsally to coxisternal seta 4c; (2) in lectotype of M. abeloosi : on both sides, with free border made up of two portions separated by curve (free border of adaxial portion arriving at cir.p on both sides, and free border of abaxial portion appearing, at right, straight and very poorly sloped in ventral projection and, at left, concave close to dis); walls equipped with small, clear and spot-like areas: one in both superior and inferior walls at right and in inferior wall at left, and two (one dorsal to other) in superior wall at left; note that longest area is area of ca. 10.0 µm long in right superior wall (roughly oval and with well discernible dots) and smallest area is upper area in left superior wall (roughly triangular and also with dots; nature of spot-like areas and dots not determined). Posterior part of discidia: not studied; note however that, in both species, as in other Brachypylina (e.g. Grandjean, 1960b: Fig. 3 View FIGURE 3 a), free border of posterior part of discidia (incompletely seen in Fig. 2 a) slopes to circumtrochanteral opening IV (i.e., more precisely, to half anterior and not sunken border of circumtrochanteral opening; cf. Grandjean, 1968: 359; 1970: 448).
Ridges associated with superior wall of discidia: (1) carinae kd ( Fig. 2 a): extending dorsally either from floor of widening o.d. of acetabular tectum IV (as in most M. grootaerti specimens and as on both sides in lectotype of M. abeloosi ) or from commissure j.d. of acetabular tectum IV to posterior part of discidium ventrally (in M. grootaerti , coming into contact with free border of posterior part of discidium, as shown in Fig. 2 a, or not; in M. abeloosi , touching free border of posterior part of discidium only at right); in both species, with dorsal part of kd being roughly parallel to and at some distance from half anterior border of circumtrochanteral opening IV; note that: (i) in some instances in M. grootaerti , o.d. is reduced by bringing j.d. nearer to border of circumtrochanteral opening and nevertheless kd appears to reach narrowed floor of o.d.; (ii) in one M. grootaerti specimen on one side, with dorsal ending portion of kd forked (anterior branch touching j.d. and posterior branch arriving at floor of o.d.); ridges associated to kd: in some M. grootaerti specimens, with companion ridge emerging from dorsal portion of kd and running to j.d.; in lectotype of M. abeloosi : at left, with auxiliary ridge ventrally extending approximately parallel to and close to kd, and dorsally deviating from kd and outwardly finishing near lower branch of ridge equivalent to regular, dorsal and posterior element associated with R in M. grootaerti (see above); no associated ridge, at right; (2) other dorsal ridges: in M. grootaerti : probably occasional; generally thin and poorly visible in LM; straight or curved; situated at some distance from dis (for example, Fig. 2 a shows one oblique companion ridge more luminous than adjacent cuticle and possibly coming into contact with kd); in lectotype of M. abeloosi : at right, with two ridges outwardly originating from free border of posterior part of discidium near dis and running parallel to free border of anterior part of discidium (then anterior element seeming to end abruptly far from cir.p whereas posterior element deviating first from free border of discidium and forming anterior limit of existent spotlike area, and then drawing near and reaching ending portion of free border of discidium near cir.p); at left, with one shorter element probably in contact with lower spot-like area, and then possibly touching cir.p.
Ridges associated with inferior wall of discidia: (1) in M. grootaerti : three to eight (most often four or five; four labelled 10 in Fig. 3 View FIGURE 3 a); posterior ridge going to and arriving at floor of ventral widening o.v. of acetabular tectum IV and ordinarily hiding largely posterior part of discidium in ventral projection (for example, in Fig. 3 View FIGURE 3 a, posterior part of discidium appears to be almost completely hidden by posterior ridge except small portion whose free border is represented by short transverse line adaxial to dis); other ridges running to and reaching (or not) ventral commissure j.v. of acetabular tectum or ventral prolongation of free border of acetabular tectum (see below) or cir.p (for example, in Fig. 3 View FIGURE 3 a, anterior ridge extends to and ends near prolongation of free border of acetabular tectum whereas both medial ridges finish at j.v.); note that: (i) ridges are frequently bent in ventral projection (yet sometimes anterior ridges or more rarely both anterior ridges appearing straight, and posterior ridges appearing somewhat sinuous); (ii) some ridges (mainly short anterior ones) probably do not arrive at inferior wall of discidium and therefore can be regarded as auxiliary elements (detailed examination not carried out); (2) in lectotype of M. abeloosi (partially studied), six ridges on both sides appearing curved in ventral projection; posterior ridges not hiding posterior part of discidium in ventral projection; at left, anterior ridge bearing coxisternal seta 4c; with auxiliary carinae located adaxially to posterior ridges (at right, composed of two elements and, at left, of one element longer and ending at some distance from anterior tooth Δ ’ of podosoma-trochanter articulation IV).
Carinae extending free border of acetabular tecta III and IV. Dorsal, oblique prolongations kf ( Fig. 2 a): in M. grootaerti : more frequently straight (as on both sides in lectotype of M. abeloosi ) than bent or sinuous; either arriving at carina kv (see above) or more often ending not far from kv (in such cases, longest distance measured longitudinally between extremity of kf and kv = 5.4 µm, and shortest distance = 1.7 µm, n = 10; in lectotype of M. abeloosi , distance kf -kv of ca. 8.5 µm on both sides); occasionally with ventral companion ridge proceeding either from intermediary part of kf or from posterior part of kf and running posteriorly and roughly parallel to kf. Dorsal, oblique prolongations kh: in M. grootaerti ; either straight or more frequently straight posteriorly and bent anteriorly over more or less long distance (as in Fig. 2 a) or curved over overall length (seldom anteriorly with second curvature directed dorsally or anteriorly); finishing at some distance from kg (see above); usually without crest line anteriorly over more or less long distance whereas crest line is always observed posteriorly; in lectotype of M. abeloosi : kh relatively short on both sides; at right, feebly bent anteriorly and with dorsal, longitudinal associated ridge (poorly perceptible and not studied); at left, straight posteriorly and, anteriorly, slightly curved dorsally.
Ventral prolongations of free border of acetabular tecta III and IV: (1) in M. grootaerti : rarely indistinguishable; extending anteriorly and obliquely to cir.p (prolongation of free border of acetabular tectum IV sometimes transverse) and coming into contact with crest line of cir.p or not; in acetabular tecta III: more frequently bent ( Fig. 3 View FIGURE 3 a) than straight (note that anterior ending portion is occasionally more or less straight and parallel to cir.p over more or less short distance; for instance, in two specimens at left, parallel ending portion ca. 6.0 µm and 8.0 µm long); more often ending posteriorly to seta 3c than approximately on transverse level with 3c or, as shown in Fig. 3 View FIGURE 3 a, more or less anteriorly to 3c; in acetabular tecta IV: more frequently straight than curved (in Fig. 3 View FIGURE 3 a, ventral prolongation is however bent); with mean length = 6.0 µm in ventral projection (range: 4.3–7.7 µm, n = 8); (2) on both sides in lectotype of M. abeloosi : ventral prolongations of free border: oblique in acetabular tecta III and IV; in acetabular tecta III, in contact with crest line of cir.p distinctly posteriorly to 3c; in acetabular tecta IV, not touching crest line of cir.p; note that ventral prolongation in IV: (i) at right, seems to be reached by anterior ridge associated to inferior wall of discidium; (ii) is ca. 3.5 µm long on both sides.
Circumpedal carinae cir.p. Anterior, longitudinal part of cir.p: in both species, showing sinuous course ( Fig. 3 View FIGURE 3 a), and being farthest from plane of symmetry on transverse level with discidium; in addition to relations with carinae cl as mentioned above, ending portion of cir.p arriving at carina ku in both species ( Fig. 3 View FIGURE 3 a; for M. abeloosi , according to lectotype and Fig. 2 c of Lions, 1978); yet, in M. grootaerti , cir.p sometimes appearing attenuated or possibly lacking over short distance posteriorly to ku; according to observations in serial sections in M. grootaerti , only limited portion of cir.p ventrally to pedotecta II appears to be laminiform (possibly also in other places in M. abeloosi ) whereas elsewhere (with exceptions indicated above) cir.p being more or less high, and with apparent outline distinctly angular in some places (in other places, apparent outline more frequently convex on one side and either straight or concave on other side) and with crest line acute or rounded ( Fig. 3 View FIGURE 3 b) in cross-sections.
Posterior, vertical part of cir. p: ending abruptly at some distance from border of ventral shield (in M. grootaerti , mean distance = 17.2 µm, range: 12.8–23.0 µm, n = 5; in lectotype of M. abeloosi , distance of ca. 11.0 µm and 14.5 µm at left and at right, respectively; in one specimen on one side, with ending portion forked); associated carinae: (1) anteroventral and oblique companion carina: arising, in M. grootaerti , just dorsally to small discontinuity observed in cir.p (taking shape of reduction or more often of curvature) and, in lectotype of M. abeloosi , ventrally to weak bend of cir.p at left (no discontinuity found at right); running to carina kh outwardly by passing along outer edge of acetabulum IV (in lateral orientation, anteroventral companion carina is projected just on apparent outline of cotyloid wall of acetabulum) and seeming to end at some distance from kh in lectotype of M. abeloosi (possibly also in M. grootaerti , yet extremity of anteroventral companion carina is hard to see in LM as well as in SEM in M. grootaerti ); (2) other anterior companion ridges: in one M. grootaerti specimen on one side, consisting of ridge more ventral than anteroventral companion carina (going to and probably finishing close to antero-ventral companion carina); in lectotype of M. abeloosi on both sides, of ridge emerging from ending portion of cir.p dorsally to junction cir.p-kl (extending obliquely to and ending far from border of ventral shield; element longer and more accentuated at right).
Other lateral features. Pleurophragmata hl. Not perforated in any of both species. Base of hl set on bottom of large sejugal depression (see above) and with apparent outline, in lateral projection, of variable form in M. grootaerti (roughly rectangular or oblong or more rarely circular) and approximately circular at left and oblong at right in lectotype of M. abeloosi . Apparent outline of hl: in dorsal projection, generally roughly triangular (as at right in Fig. 1 View FIGURE 1 and on both sides in Fig. 1 View FIGURE 1 of Lions, 1978) and more seldom coarsely rectangular or with extremity broadened and more or less rounded (extremity of hl well rounded at left in Fig. 1 View FIGURE 1 ); roughly crook-shaped according to serial sections transverse through pleural region and medio-longitudinal through hl. Occasionally, pleurophragma weakly bent anteriorly (in dorsal projection in lectotype of M. abeloosi , appearing slightly curved anteriorly and acuminate at left and straight and with broad and rounded extremity at right) and more often bearing one point (left side in Fig. 1 View FIGURE 1 and also in lectotype) than two points (as probably at right in lectotype, yet confirmation is required because points are not very distinguishable) or none (at right in Fig. 1 View FIGURE 1 ).
Latero-dorsal dotted areas. Located on both sides posteriorly to pleurophragma hl and ventrally to carina kl. In M. grootaerti , of variable size and form, and ordinarily with blurred limits. In lectotype of M. abeloosi , well rounded at left and with clear limits on both sides. Observations in optical, cross-sections indicate that, in M. grootaerti , dots could correspond to porose canals rather than microsculpture. Latero-dorsal dotted areas would then be analogous to sub-alar areae porosae Ah known in brachypyline Poronota ( Grandjean, 1962a: 396; note that Poronota are not closely related to Microzetidae ), at least if laterodorsal dotted areas communicate with dermal glands as do areae porosae Ah and other areae porosae in Poronota (cf. Alberti & Coons, 1999: 711). Confirmation regarding existence of dermal glands connected with laterodorsal dotted areas is required by observations in TEM.
If dermal glands are not detected, latero-dorsal areas would then represent zones with normal cuticular pores more densely distributed (note that Grandjean, 1956a:142, hypothesizes that three sub-alar areas in poronotic Pilogalumna ornatula Grandjean, 1956 , are zones with high density of normal cuticular pores).
Podocephalic fissures F. Anterior transverse part of F, in ventral projection: in M. grootaerti , more frequently straight ( Fig. 3 View FIGURE 3 a) than bent; in lectotype of M. abeloosi , curved on both sides (as seen in Fig. 2 c of Lions, 1978). Posterior longitudinal part of F: in both species, extending to ventral part of circumtrochanteral openings I and, consequently, hidden posteriorly by pedotectum I in lateral and ventral ( Fig. 3 View FIGURE 3 a) projections; in M. grootaerti , running at short distance from and roughly parallel or sometimes definitely parallel (as shown in Fig. 3 View FIGURE 3 a) to posterior part of laterorostral carina lr; in lectotype of M. abeloosi , on both sides, appearing in lateral projection to be overhung by lr (at left, outwardly closer to and parallel to lr). Hidden part of podocephalic fissures (as well as supracoxal spines eI) not studied.
Muscle impressions posterior to acetabulum IV. In M. grootaerti , made up of dozen impressions with thick dots in some places (nature of dots not determined) and more frequently forming single, elongated and oblique group than two imperfectly separated groups (in lectotype of M. abeloosi , one single group similar to groups in M. grootaerti is found on both sides, and thick dots appear more numerous at left). In M. grootaerti , muscle impressions usually situated horizontal level with junction between cir.p and anteroventral and oblique companion carina of cir.p (mentioned above) whereas, in lectotype of M. abeloosi , dorsal muscle impressions at left and medial muscle impressions at right appearing to be level with junction cir.p -carina. Note that: (i) muscle impressions take place of small tendinous insertions γ observed in most Brachypylina in vicinity of cir.p ( Grandjean, 1957a: 155); (ii) just like insertions γ in some Brachypilina (e.g. Grandjean, 1960b: 116), most of muscle impressions are projected at some distance from cir.p in ventral view and closer to and even upon cir.p in lateral view.
Ventral region of podosoma ( Fig. 3 View FIGURE 3 a). Relatively to ventral shield, ventral region of podosoma appearing narrower than in some other Brachypylina as e.g. Autogneta penicillum Grandjean, 1960 , in which ventral region of podosoma is regarded as large ( Grandjean, 1960a: 351); for instance, breadth of ventral region of podosoma (measured between anteroventral extremities of pedotecta I)/breadth of ventral shield (measured just posteriorly to genital valves) ratio is ca. 0.48 in M. grootaerti (n = 2), 0.54 in M. abeloosi ( Fig. 2 c of Lions, 1978) and 0.65 in A. penicillum ( Fig. 1 View FIGURE 1 b of Grandjean, 1960a).
Remarkable traits. Lateral, V-shaped carinae cv of mentotectum (lacking in M. abeloosi but other carinae are present as indicated below). Adaxial branch: accentuated and frequently roughly straight; anteriorly, infrequently reaching free border bm of mentotectum and, posteriorly, often ending at posterior limit of mentotectum (and even arriving and finishing at epimeron 1). Abaxial branch: frequently thinner and curved; arising from posterior part of adaxial branch and going to carina cl (see above). Associated ridges: sometimes with two ridges comprising companion, additional element emerging from posterior part (or extremity) of adaxial branch (so that cv appears tridentate) and companion or auxiliary element found adaxially to companion, additional ridge; in few instances, with only auxiliary, longitudinal ridge adaxial to cv ( Fig. 3 View FIGURE 3 a). Note that presence of cv and cl makes difficult limit of mentotectum to be studied in anteroventral region of podosoma in LM; yet observations in SEM indicate that limit of mentotectum could be situated either between cl and abaxial branch of cv (and then abaxial branch of cv would run mainly onto inferior wall of mentotectum) or between both branches of cv (and then abaxial branch of cv would extend onto epimeron 1).
In lectotype of M. abeloosi , with two carinae on both sides, namely: (1) lateral and oblique carina (ca. 16.0 µm long on both sides): abaxially, bent and running anteriorly over some distance (at left terminating not far from extremity of cl whereas at right ending portion rounds before touching extremity of cl); adaxially, finishing abruptly (at left, curved anteriorly and with short ending portion beyond curve); (2) adaxial, associated carina made up: at left, of long element appearing as extension of previous, lateral carina towards plane of symmetry and probably ending close to plane of symmetry (both carinae are not in contact, yet extremity of associated carina is located just posteriorly to adaxial ending portion of lateral carina); at right, of distinctly shorter ridge parallel to ending portion of lateral carina (outwardly accompanied with thin and very short ridge extending from ending portion of lateral carina up to nearly posterior extremity of short ridge).
Long, lateral gutters (seen in cross-section in Fig. 3 View FIGURE 3 b; not studied in lectotype of M. abeloosi ). Running along anterior, longitudinal part of cir.p. According to observations in SEM, appearing less marked anteriorly to pedotectum II (or even absent in places, according to some observations in serial sections). With adaxial limit bg not very visible or even indistinguishable in ventral projection in LM (represented over some distance in Fig. 3 View FIGURE 3 a).
Epimeral furrows s4. In M. grootaerti , consisting of two parts on both sides: (1) lateral (abaxial) part (represented by line in Fig. 3 View FIGURE 3 a): narrow (distance between borders <1 µm according to observations in SEM) and probably shallow (depth not measured); usually making large, transverse curve concave anteriorly while extending between lateral gutter of podosoma (which s4 reaches or not) and ventral tendinous apophysys β (note that, in ventral orientation, β is projected either adaxially to extremity of lateral part of s4 or more often upon or nearly on same longitudinal line as extremity of lateral part of s4); (2) adaxial part less perceptible in ventral projection in LM (and some portions of anterior border occasionally even hard to detect); first broadening out when coming close to plane of symmetry ( Fig. 3 View FIGURE 3 a) and then passing just anteriorly to inclined rim rtg of anterior border of circumgenital opening so that appearing to form large curve convex anteriorly with s4 of other side. In lectotype of M. abeloosi , s4 seeming to exist only laterally and appearing appreciably less distinguishable than in M. grootaerti in ventral projection.
Mentotectum. In both species, with apparent longitudinal outline curved anteriorly in plane of symmetry. Maximum height (measured in lateral orientation, in or nearly in plane of symmetry, from line fitting to surface of epimeron 1): in M. grootaerti , mean value = 12.9 µm (range: 11.1–14.3 µm, n = 6); in M. abeloosii , ca. 8.0 µm according to Fig. 3 View FIGURE 3 a of Lions (1978) and to lectotype. In lectotype of M. abeloosi , with unpaired, sagittal and roughly rhombic area: anterior and lateral parts of area darker than surroundig cuticle; posterior, longer part dotted and encroaching posteriorly upon epimeron 1 (nature of area and dots not determined).
Epimeral elements. (cf. Grandjean, 1952). Epimeral elements 3 lacking in both species. Epimeral, thickened borders. Border bo1: in both species, without discernible adjoining muscle impressions and with rather comparable breadth in vicinity of plane of symmetry; in M. grootaerti , anterior limit of bo1 sometimes not or very poorly visible in ventral projection so that, as shown in Fig. 3 View FIGURE 3 a, only two lines are seen posteriorly to bm in depth (i.e. line ba.m corresponding to base of mentotectum and line b1 representing posterior limit of bo1) instead of three lines as in lectotype of M. abeloosi . Border bo2: in both species, not arriving at plane of symmetry contrary to other epimeral borders; in lectotype of M. abeloosi , instead of being perpendicular to plane of symmetry as in M. grootaerti ( Fig. 3 View FIGURE 3 a), directed obliquely and anteriorly on both sides, at angle somewhat smaller than 90 degrees.
Sejugal border bo.sj: (1) lateral parts (i.e. parts on which sejugal half apodeme ap.sj is fixed): in both species, also inclined anteriorly and forming clearly acute angle relative to plane of symmetry; generally in M. grootaerti and in lectotype of M. abeloosi , with narrowed portion perceptible in ventral projection on level with adaxial part of sejugal apodeme; (2) with transverse ridges on internal surface of cuticle, namely: in epimeron 2, on both sides in M. grootaerti , usually one ridge running abaxially from narrowed portion of bo.sj (more frequently from abaxial limit of narrowed portion) to middle part of bo.sj at some distance from plane of symmetry (not represented in Figs 3 View FIGURE 3 a; not detected in lectotype of M. abeloosi ); in middle part of bo.sj, ordinarily one and seldom two or three internal ridges on both sides in M. grootaerti , and one ridge at right and possibly three ridges at left in lectotype of M. abeloosi .
Border bo4: in both species, according to optical sections transverse through pleural region and longitudinal through bo4, maximum height detected close to plane of symmetry (a common trait in Microzetidae ; cf. Grandjean, 1936a: 62); with some transverse striae-like elements on internal surface in middle region of bo4, poorly distinguishable in M. grootaerti (not shown in Fig. 3 View FIGURE 3 a; note that, in lectotype of M. abeloosi , very narrow zone of cuticle appears clearer along anterior internal ridge of bo4). Sternal border: in both species, developed only in middle region of metapodosoma lengthwise; occasional in M. grootaerti (not seen in Fig. 3 View FIGURE 3 a) and frequently appearing broad over overall length and generally directed obliquely; in lectotype of M. abeloosi : with intermediary part relatively narrow; broadening out over longer distance anteriorly than posteriorly.
Furrows 2 and sejugal furrows sj. In ventral orientation, hard to detect in LM in both species. In lateral orientation, both furrows appearing shallow and s.sj deeper in both species whereas s2 seeming to be obliterated in M. grootaerti and very weak in M. abeloosi , in vicinity of plane of symmetry.
Apodemes. With three apodemes, namely: ap.1, ap.2 and ap.sj; each apodeme consisting of two half apodemes which are not perforated; ap.1 and ap.2 with approximatively similar maximum height which is less in ap.sj. Half ap.1 and ap.2 not studied in detail in both species; yet note that: (i) ‘point’ of half ap.1 ( Grandjean, 1952) is sometimes not perceptible in ventral projection in M. grootaerti (yet represented in Fig. 3 View FIGURE 3 a) and is observed on both sides in lectotype of M. abeloosi (as in Fig. 2 c of Lions, 1978); (ii) in both species, small protuberances are detected in lower part of anterior wall in half ap.2. Half sejugal apodemes ap.sj (not studied in lectotype of M. abeloosi ): roughly trapezoidal in apical projection ( Fig. 3 View FIGURE 3 b); dorsoadaxial region lobe-shaped and weakly inclined posteriorly; part of adaxial border between lobe and base of half apodemes ordinarily curved anteriorly (in Fig. 3 View FIGURE 3 b, line seen on adaxial side and appearing convex laterally represents schematically free border of half apodeme, and line adaxial to previous one corresponds to bend of free border; note that degree of curvature and breadth of bent portion are variable); with reticulation made up of rounded, irregular, hardly distinguishable and poorly raised meshes (possibly borne by posterior wall of apodeme, yet confirmation is required); with ridge extending obliquely and dorsally onto ventroadaxial region of anterior wall of apodeme (well discernible in serial sections; not represented in Fig. 3 View FIGURE 3 b).
Coxisternal setae. Resembling M. abeloosi in number (formula: 3-1-3-3), in smooth aspect, and in location of setae 3c and 4c. In M. grootaerti , setae 3a more frequently inserted on epimeral border bo.sj than posterior to bo.sj as on both sides in lectotype of M. abeloosi . In two M. grootaerti specimens, with one additional seta between setae 4a and 4b whereas, contrary to lectotype of M. abeloosi at left (as reported above), no doubling of seta 3c found.
Tracheal system (not studied in detail in M. abeloosi ). Location of sejugal stigmata st anterior to base of pedotectum II confirmed in Fig. 3 View FIGURE 3 b; however, position of st relative to common extremity of kpII and free border of acetabulum II (see above) could not be ascertained because observation is hindered by pedotectum II and carinae associated to inferior wall of pedotectum II (attempts to detect position of st in lectotype of M. abeloosi unsuccessful, as were such attempts by Lions, 1978: 411). Branching of tracheae I situated close to stigma, with anterior branch (tr.1a) appearing to originate from posterior branch (tr.1p) (disposition frequently occuring in Brachypylina and called ‘branching with turning back’ by Grandjean, 1968: 378), and with posterior branch as usual longer (and moreover, curved in prosoma). In sejugal tracheae, as shown in Fig. 3 View FIGURE 3 b, with common trunk between vestibulum ve (which is distinctly bent and as usual enclosed in half sejugal apodemes ap.sj) and branching (note that common trunk is either lacking or very short in many Brachypilina; cf. Grandjean, 1934: 127), and with both branches (tr.sj. α) and (tr.sj. π) entering opisthosoma (abaxial branch tr.sj. α is longer and generally twisted). Tracheae III long, bent in posterior region of opisthosoma, and occasionally twisted in dorsal region of opisthosoma.
Ventral region of opisthosoma. Distance between circumgenital and circumanal openings: mean value = 25.5 µm in M. grootaerti (range: 23.7–28.0 µm, n = 6); ca. 22.0 µm in M. abeloosi according to lectotype and to specimen represented in Fig. 2 c of Lions (1978). Both circumgenital and circumanal openings appearing relatively larger in M. grootaerti . Observations in serial sections in M. grootaerti reveal that, anteriorly, border of ventral shield is deviated abaxially and internal border thickening ne is outwardly absent whereas, posteriorly, no deviation of border is observed and ne is present (accordingly, border zone of ventral shield, when cut crosswise, appears broadly as in Fig. 2 a of Grandjean, 1959c).
Remarkable traits. Chaetotaxy of laterodorsal lobes of vulva singular since one of four eugenital setae regularly detected in Oribatida (labelled τ a, τ b, τ c and τ 1; cf. Grandjean, 1956b: 102) is lost in M. grootaerti ( Fig. 4 View FIGURE 4 a) and in two M. abeloosi specimens studied by Lions (1978: Fig. 3 View FIGURE 3 c); note that: (i) suppressed seta is assumed to be τ a; (ii) in M. grootaerti , setae are at each corner of flatter triangle, nearly lined up in lateral projection. Distal setae τ 1: base with canal clearly perceptible in M. grootaerti (canal hard to see in setae τ b and τ c); not hook-like but slightly sinuous in M. abeloosi (according to Fig. 3 View FIGURE 3 f of Lions, 1978); tapering off distally in both species.
Genital region. Genital valves. In M. grootaerti , lock coaptation system involving one (as seen in Fig. 2 d of Lions, 1978, in M. abeloosi ) or more frequently two elements short (maximum length of ca. 7.0 µm in one specimen on one side) and situated near each other posteriorly on adaxial border of each genital valve (note that elements are often clearly distinguishable, with apparent outline curved or roughly angular in ventral projection).
Vulva. In both species: as in other Brachypylina (e.g. Behan-Pelletier, 1991), anterior genital papillae Va situated further from surface of genital valves than genital papillae Vm and Vp when valves are closed and seen in ventral projection; as usual, six coronal setae present; with fringe at distal border of vulva, made up of small, conical elements ( Fig. 4 View FIGURE 4 a). In M. grootaerti : papillae Va ( Fig. 4 View FIGURE 4 b) cone-shaped, as long as broad, rounded apically and distinctly shorter than papillae Vm and Vp which have usual form (Va not studied in M. abeloosi ); longitudinal folds of external wall without granules as shown in M. abeloosi ( Fig. 3 View FIGURE 3 c of Lions, 1978); setae ψ 1 of ventral lobe: appearing very short in M. grootaerti (mean length = 3.4 µm, range: 3.3–3.5 µm, n = 5; length of 7.8 µm in M. abeloosi according to Fig. 3 View FIGURE 3 c of Lions, 1978); lost in one specimen on one side.
Anal region. In both species: preanal organ comparable ( Figs 3 View FIGURE 3 b and 3e of Lions, 1978); adanal lyrifissures iad located posteriorly to adanal setae ad3 and close and more or less parallel to lateral border of circumanal opening; with low, blunt, longitudinal or oblique ridges in vicinity of adanal setae ad2: (1) in M. grootaerti , sometimes with one or more rarely with two ridges on one side; for example, with two oblique ridges at right in one specimen, one anterior to ad2 (extending to and ending at some distance from lyrifissure iad) and one posterior to ad2 (finishing abruptly far from adanal seta ad1); (2) in lectotype of M. abeloosi , with two longitudinal ridges abaxial to and at some distance from seta ad2 at right and none at left; with transversal, padlike protuberance posterior to anal valves (i.e. small ‘rump’ as in many other Oribatida ), bearing adanal setae ad1, and clearly discernible in lateral projection. In M. grootaerti , with shallow and transverse concavity frequently detected in both anterior and posterior parts of anal valves or, if not, either found only in posterior part of valves or outwardly lacking.
Surface setae. No difference between both species regard to number (as in many brachypyline Oribatida , 12 pairs of setae present: six pairs of genital setae disposed in two rows comprising two setae abaxially and four setae adaxially; one pair of aggenital setae; three pairs of adanal setae; two pairs of anal setae), to smooth aspect (except anterior genital setae in M. grootaerti , which are barbed and also longer than other genital setae) and to location of setae ad3 (as usual posterior to anterior border of circumanal opening). Following variations observed in M. grootaerti (12 specimens surveyed): (1) presence of five genital setae on one side due to loss of one seta either in adaxial row (in two specimens) or in abaxial row (in one specimen); (2) variability in position mainly in both abaxial genital setae; (3) presence of three anal setae lined up on one side in three specimens.
Gnathosoma, chelicerae and pedipalps. Remarkable traits. Rutellar microtube (4 in Fig. 2 b). Built up by both anterior, ending parts of rutella, each part forming half of microtube, from unmodified rutella (which would correspond to Fig. 6a of Grandjean, 1957b), by drawing nearer to plane of symmetry posteriorly and by bending to plane of symmetry medially and anteriorly. Each half microtube: appearing to be thin, membranous, with lobeshaped ventral free border (in lateral projection) as in M. abeloosi (labelled 4 in Figs 4 View FIGURE 4 c and 4d of Lions, 1978); bearing anterodorsal, long, flat and unciform element 1 (directed anteriorly and abaxially; much shorter, simple and point-like in M. abeloosi ) and posteroventral and small point 6 (directed anteriorly and adaxially; in Fig. 2 b, extremity of right point is seen ventrally to left point; apparently without equivalent in M. abeloosi ). Moreover, dorsal end of rutellar microtube appearing to be incompletely obtured by elements 5 (in Fig. 2 b, left element 5 seems to be partially hidden by right element, as shown in three other specimens; elements not studied in details; not mentioned in M. abeloosi ). Note that: (i) point-like and unciform elements 1 appear to be very uncommon traits in Brachypylina since elements 1 are usually tooth-like (e.g. Grandjean, 1962b: 91); moreover, to date, five microzetid species are known to possess point-like elements 1 (according to observations reported by Lions, 1978: 413); (ii) elements 2 and 3 in M. abeloosi and combs are outwardly lacking in M. grootaerti ; (iii) in specimens treated with lactic acid (even cold), both anterior parts of rutella are separated so that no microtube is detected (accordingly, Lions who used exclusively preparations in lactic acid for description of M. abeloosi could not have observed rutellar microtube).
Oral setae. Bifurcated in M. grootaerti ( Fig. 2 b) whereas four oral setae appear to be simple phaneres in M. abeloosi ( Fig. 4 View FIGURE 4 a of Lions, 1978).
Subcapitulum. (cf. Grandjean, 1957b and 1957c). Additional considerations on rutella. Atelebasic (cf. Grandjean, 1957b) in both species. Dorsal side of manubrial zone with dots (appearing to correspond to pore canals rather than microsculpture) extending posteriorly onto large carina gc up to base of pedipalps (with comparable dotted area in lectotype of M. abeloosi ).
Carinae of coaptation with camerostome. In both species, genal carinae cog reaching collar of rutella ( Fig. 2 b). Latero-dorsal carinae coh: (1) in M. grootaerti : straight, curved or weakly sinuous; running from posterolateral angle of mentum to labiogenal suture lg and finishing at some distance from lg or infrequently not far from lg; with associated carinae consisting of dorsal auxiliary ridge (anteriorly more or less parallel to coh and posteriorly deviating from coh) and, occasionally, of ventral ridge (either longitudinal and roughly parallel to coh or more often transverse, in contact with coh or not, straight and ending abruptly at some distance from coh); (2) main differences in lectotype of M. abeloosi : on both sides, coh divided into two branches anteriorly (on both sides, dorsal branch extending to dorsal auxiliary ridge and, at left, possibly arriving at dorsal auxiliary ridge; at left, ventral branch longer and running to and not touching lg); with two dorsal auxiliary ridges close together, at right, and with one dorsal auxiliary ridge forked posteriorly, at left; on both sides, with oblique and curved, ventral companion ridge seeming to reach boundary of narrow and weakly sloping up area along posterior border of mentum (similar area also detected in M. grootaerti , yet best seen from posterior view on a separated mentum; note that this area might be used for coaptation with mentotectum).
Other features. Similarities: diarthry; unpaired, inferior commissural induration ic of mentum more or less rounded (as in lectotype of M. abeloosi ) or oval in ventral projection (in M. grootaerti , with two small spots or less frequently with two additional short points anteriorly); adaxial side of large carinae gc with narrow, elongated and not dotted area in contact with manubrial articulation and consequently possibly corresponding to fissure-like part of manubrial articulation (confirmation is however required since area appears to be directed obliquely and posteriorly in dorsal projection). Dissimilarities: in M. grootaerti , anterior region of mentum weakly depressed transversally posteriorly to lg (not indicated in M. abeloosi ), and dorsal internal nervures ne longer ( Grandjean, 1957c: 90).
Traits not studied in M. abeloosi : (1) labrum with two distinct, longitudinal lines fairly parallel over long distance in dorsal projection, yet drawing nearer anteriorly and frequently diverging posteriorly approximately from level with upper commissures Js (according to observations in serial sections, lines seeming to correspond to lateral limits of dorsal surface of labrum which appears convex or flat in cross-section); (2) porosity distinguishable in bottom of cheliceral grooves; (3) subcapitular foramina well perceptible and ducti of subcapitular glands sometimes detectable; (4) median notch of capitular apodeme relatively short (at least by comparison with notch represented in Xenillus clypeator Robineau-Desvoidy, 1839 , in Fig. 1 View FIGURE 1 of Grandjean, 1957c).
Setae. In M. grootaerti , unlike M. abeloosi , medial sucapitular setae m barbed whereas, in both species, anterior genal setae a and hysterostomatic setae h being smooth. Laterocoxal setae e straight as in lectotype of M. abeloosi .
Chelicerae. According to observations in serial sections in M. grootaerti , distal portion of cheliceral bodies with shallow concavity on both sides and with weak convexity ventrally on abaxial side (not studied in detail; ventro-abaxial convexity not apparent in two preparations). Similarities. Chelicerae regarded as not elongated (mean height/length ratio = 0.39, range: 0.37–0.40, n = 9, in M. grootaerti and ratio of 0.41 in keeping with Fig. 4 View FIGURE 4 b of Lions, 1978, in M. abeloosi ) seeing that, according to Grandjean (1964a: 705 and Fig. 3 View FIGURE 3 b), ratio of 0.36 corresponds to chelicerae ‘a bit more elongated than on average in Brachypylina’. Trägårdh’s organs: in M. grootaerti , observations in serial sections reveal that Trägårdh’s organs do not project beyond apparent, dorsal outline of cheliceral body in lateral projection (as likewise in M. abeloosi ) and appear to be roughly quadrangular in cross-section over nearly whole length (not stated definitely in M. abeloosi ). Setae cha and chb: with comparable location and form (cha bent distally; chb straight); length: cha: ca. 29.0 µm in M. grootaerti and ca. 27.5 µm in M. abeloosi ; chb: ca. 14.5 µm in M. grootaerti and 13.0 µm in M. abeloosi (measured in two specimens in M. grootaerti and according to Fig. 4 View FIGURE 4 b of Lions, 1978).
Dissimilarities. Following features in M. grootaerti to be compared with traits in Fig. 4 View FIGURE 4 b of Lions (1978): (1) dorsal horn of fixed digits: not always straight but occasionally poorly curved ventrally; (2) movable digits: with three teeth (proximal tooth appears as small, ordinary tooth except in few instances where proximal tooth is modified into rounded and poorly protruding element) and often with little or very little, proximal protuberance (which could be regarded as vestige of tooth labelled 4i in general system of notation used by Grandjean, 1947: 336); (3) adaxial fossae fc: with closed, oval or approximately triangular outline, and with some more or less discernible ridges arising from border of fossae. In addition, whereas only dots of unknown nature are found here and there in M. abeloosi (according to Lions, 1978: 413), presence of two areae porosae in M. grootaerti : dorsoabaxial area porosa extending onto dorsal side (from fixed digit distally to either seta cha or less frequently line en; cf. Grandjean, 1947) and onto abaxial side up on level with seta chb (note that pores of abaxial side are sometimes difficult to see); ventral area porosa covering whole ventral region of cheliceral bodies posteriorly to movable digit (including posterior extension of ventral region on which tendon tvi is fastened; Grandjean, 1947).
Pedipalps. In both species (according to Fig. 4 View FIGURE 4 g of Lions, 1978, in M. abeloosi ): (1) lengthening and cambering of tarsi; (2) eupathidia: subultimal eupathidia su ζ longer than others; ultimal eupathidia ul ζ ’ and ul ζ ’’ remote from each other, ul’’ ζ being highest and brought nearer anteroculminal eupathidia acm ζ (probably somewhat less markedly in M. grootaerti ); note that: (i) canal of su ζ is well visible in high magnification in LM whereas canal of ul ζ ’ and ul ζ ’’ is barely detectable; (ii) though presence of canal cannot be evidenced, acm ( Fig. 3 View FIGURE 3 c) is assumed to be eupathidium because of similitude of form to other eupathidial setae; (3) setae: usual formula: 0-2-1-3-9; in genua, with seta inserted on abaxial side; in tarsi, with double prime disjunction in pair of lateral setae lt and in pair of ventral setae vt, and with prime basculation in (ul ζ). In M. grootaerti : (1) femora with two ridges (not mentioned in M. abeloosi ), ventral ridge straight and located adaxially to seta v’’ and adaxial ridge bent; (2) in tarsi: lt’ inserted more distally and approximately on vertical level with vt’; as shown in Fig. 3 View FIGURE 3 c, free solenidia ω shorter and inserted appreciably more distally (note that solenidia are laid on cuticle and thereby are poorly perceptible in lateral projection).
Legs. Remarkable traits. Internal reticulation. In M. grootaerti , detected in all segments except in trochanters I and II, genua, tarsi I and II (but see below), and ambulacra. Meshes: with irregular form and size (mean length = 7.0 µm, range: 4.5–11.0 µm, n = 30); in femora IV, usually well apparent proximally on adaxial side due to broadened borders; in tibiae IV, frequently making large and roughly oval area approximately in intermediary part of abaxial side; in tarsi III and IV, hard to see on ventral side and generally poorly distinguishable elsewhere except often one proximal mesh both on dorsal side in III and on abaxial side in IV. Proximal, dull spot frequently found on dorsal side in tarsi II and another spot less often observed on abaxial side in tarsi I might be isolated meshes. In lectotype of M. abeloosi , although survey of all segments is lacking, internal reticulation is seen in places in trochanters IV, femora III and IV, and tibiae III.
Additional seta on femora I. With five instead of four setae as in M. abeloosi due to presence of ventral seta ordinarily showing median or nearly median position. Additional seta might be posteroventral seta v’’ of telofemoral verticil (which has shifted secondarily in ventral direction) since, in Brachypylina , setae v’’ in femora I appear to offer greater resistance to suppression than anteroventral setae v’ (see examples in Grandjean, 1960a: Fig. 4 View FIGURE 4 a, 1961b: Figs 3 View FIGURE 3 b and 12a, 1964b: Fig. 4 View FIGURE 4 a, among others). Additional seta is inserted either on longitudinal part of L-carina (see below) or more frequently adaxially close to L-carina.
Proral setae (p). In M. grootaerti : in tarsi I, not very different from setae (p) in M. abeloosi ; in tarsi II, III and IV, poorly detectable in LM even with high magnification, and with shape and size as in seta p’ in tarsus IV shown in Fig. 2 c.
Main similarities. Lengthening of segments. Difference in lengthening lower than 3% between both species for most segments (as well as for tarsi I and IV in M. senensis according to Figs 3 View FIGURE 3 a and 3b of Bernini, 1975), yet some segments appearing to be more elongated in M. grootaerti , especially trochanters III and IV (height/length of bulb ratio: in trochanters III, averaging 0.81, range: 0.80–0.83, n = 4, in M. grootaerti vs 0.91 and 0.94 in lectotype of M. abeloosi and 0.92 according to Fig. 6a of Lions, 1978; in trochanters IV, in M. grootaerti , averaging 0.88, range: 0.87–0.90, n = 4, in M. grootaerti vs 0.98 and 1.02 in lectotype of M. abeloosi and 1.03 according to Fig. 4 View FIGURE 4 f of Lions, 1978).
Carinae on femora I. L-shaped carina (consisting of two parts, namely transverse and longitudinal parts usually connected according to an obtuse angle): (1) transverse, adaxial part: generally largely curved in proximal direction (not seen in Fig. 1 View FIGURE 1 ), ending at dorsal side, and with proximal associated ridge more or less transverse and sometimes split into two dorsally (comparable in lectotype of M. abeloosi except for absence of associated ridge); (2) longitudinal, ventral part (not discernible in Fig. 2 a, yet well perceptible in ventral projection in SEM and also in LM) ordinarily slightly deviating distally in abaxial direction and, in addition to v’’ (as indicated above), bearing posterolateral l’’ usually at distal extremity and less often basifemoral seta bv’’ (otherwise bv’’ is inserted adaxially close to longitudinal part); with concavity developed just abaxially to longitudinal part (not studied in detail; dorsal boundary of concavity seeming to correspond to line largely curved in dorsal direction and detected on ventral and abaxial sides of femur I in Fig. 2 a); in lectotype of M. abeloosi : on both sides, longitudinal part of L-shaped carina finishing distally to l’’; bv’’ inserted on longitudinal part on both sides; l’’ fixed on longitudinal part, at right, and abaxially to longitudinal part, at left; at right, with curved associated ridge abaxially to bv’’.
Longitudinal, abaxial carina (not shown in Fig. 1 View FIGURE 1 and hard to see in Fig. 2 a): sinuous in both species (yet less markedly in lectotype of M. abeloosi ); in M. grootaerti , generally less long than in lectotype (yet clearly longer than in Fig. 5a of Lions, 1978), and with some, mainly ventral associated (companion or auxiliary) ridges (not studied in detail, yet note presence of ventral, transversal ridge extending to and reaching angle made by L-shaped carina; without associated ridge in lectotype of M. abeloosi ). Although drawing nearer to each other proximally, L-shaped carina and longitudinal, abaxial carina don’t seem to come into contact or to have common extremity.
Ventral laminae in trochanters and femora III and IV (in Fig. 3 View FIGURE 3 a, lamina is seen in optical cross-section distally in trochanter III and proximally in trochanter IV whereas small, proximal part of lamina is perceived obliquely in femur III). In trochanters IV, breadth of lamina less marked in proximal part in M. grootaerti and in intermediary part in M. abeloosi . In M. grootaerti , unlike M. abeloosi , lamina reduced to carina proximally in trochanters III ( Fig. 3 View FIGURE 3 a; note that interruption occasionally occurs between laminiform and careniform parts) and distally in femora III. Moreover, as in many other Brachypylina ( Grandjean, 1962b: 94) , with shoulder overhanging dorsally and abaxially stalk of femora III and IV ( Fig. 2 a) and prolonged by ventral lamina (in Fig. 3 View FIGURE 3 a, free border of shoulder is partially seen and labelled ep).
Transverse furrow in bulb of genua I and II. In M. grootaerti : in genua I, furrow better visible on dorsal side, and sometimes outwardly not detectable on ventral side; in genua II, furrow frequently spread on all sides (if not, then distinguishable only on dorsal and adaxial sides). In M. abeloosi : furrow found on dorsal and abaxial sides in genua I in lectotype, possibly on dorsal side in genua II (according to Fig. 5b of Lions, 1978), and on adaxial and ventral sides in genua III in lectotype; furrow not observed in genua IV.
Lyrifissures of tarsi. In both species, double prime basculation of lyrifissure in tarsi I and II, and prime basculation in tarsi III; in M. grootaerti , prime basculation in tarsi IV (in M. abeloosi , no possibility of studying lyrifissure in tarsi IV in Fig. 6b of Lions, 1978).
Porose areas. On both sides, six porose areas present, with placement either normal (according to Grandjean, 1953a: 126), i.e. adaxial in trochanters IV (yet more dorsal in M. grootaerti ) and dorsoadaxial in femora I (incompletely represented in Fig. 1 View FIGURE 1 ), II, III and IV or unusual, i.e. dorsoadaxial in trochanters III in M. grootaerti and adaxial in femora IV in M. abeloosi (as shown in Fig. 4 View FIGURE 4 e of Lions, 1978).
Main dissimilarities. Proximal depressions in bulb of trochanters III. In M. grootaerti , proximal region of bulb in trochanters III depressed either both dorsally and ventrally (so that, as shown in Fig. 2 a, bulb appearing to possess strong proximal protuberance in lateral orientation) or only dorsally (no depression detected in one individual on one side). Most likely less frequent in M. abeloosi since depressions are not represented in Fig. 6a of Lions (1978) and are not found in right trochanter III in lectotype (yet dorsoproximal depression present at left in lectotype). Note that additional observations are required to verify whether weak concavity of abaxial apparent outline seen in bulb in Fig. 3 View FIGURE 3 a (frequency not determined) arrives at dorsal and ventral depressions or not.
Concavities in bulb of femora II, III and IV. In M. grootaerti : (1) in femora II, with two dorsal concavities, namely often one in intermediary part of bulb (l’’ inserted more or less close to or less frequently on distal boundary of concavity; concavity ordinarily elongated lengthwise rather than broadwise as shown in Fig. 2 a; note that, outside concavity, dorsal apparent outline of bulb appearing flattened in lateral projection) and infrequently another concavity more proximal and less large (not evidenced in M. abeloosi ); (2) in femora III, occasionally with small, dorso-proximal concavity (refilled with dirt in Fig. 2 a; not mentioned in M. abeloosi ); (3) in femora IV, without distoabaxial concavity unlike M. abeloosi lectotype’s left side.
Carinae on femora II. U-shaped carina: (1) in M. grootaerti , with adaxial and abaxial branches appearing to near each other distally on ventral side (while running to and terminating close to v’’) when branches are seen in dorsal orientation by transparency; (2) adaxial branch (not indicated in M. abeloosi ) often sinuous and with distal portion frequently escorted by roughly parallel auxiliary ridge ( Fig. 3 View FIGURE 3 a); (3) abaxial branch (partially shown in Fig. 2 a): proximal portion forming large and transverse bend sometimes interrupted over short distance (as represented in M. abeloosi in Fig. 5b of Lions, 1978); unlike M. abeloosi , not bearing bv’’ (insertion of bv’’ being approximately in plane of pseudosymmetry in M. grootaerti ); distal portion (labelled cf in Fig. 3 View FIGURE 3 a) often with transverse, companion ridge going to and ending near bv’’ ( Fig. 3 View FIGURE 3 a). Ventroproximal and longitudinal carina: ca. 10.5 µm long (according to one specimen on one side); extending from stalk proximally to bv’’ distally, and finishing not far from bv’’ (frequency not estimated; not cited in M. abeloosi ; not seen in Fig. 3 View FIGURE 3 a).
Proximal, transverse carina in bulb of tibiae (lacking in M. abeloosi ). In tibiae I, carina detected on dorsal and abaxial sides and less frequently on adaxial side, and not observed on ventral side. In tibiae II, spread on all sides and usually better discernible on dorsal and abaxial sides. In tibiae III, often going round segment, yet occasionally not visible on adaxial and ventral sides; in tibiae IV, indistinguishable on adaxial side, rarely perceptible on adaxial and ventral sides, and generally well apparent on dorsal side. In addition, on abaxial side in tibiae I, III and IV, frequently curved (or possibly bent at angle) and sometimes with one or two diversely orientated, associated ridges (for example, in one individual on one side in tibiae III, with transverse ridge running between both sides of curve).
Ungues. In M. grootaerti , ungues III longer than ungues II whereas, according to Figs 5 and 6 of Lions (1978), length of ungues III and IV appearing fairly similar in M. abeloosi (note however that difference in length between ungues III and II can be small in M. grootaerti as illustrated in second example below). In other respects, ungues IV longer than other ungues (as in M. abeloosi ), and ungues II either longer (or slightly longer) than ungues I (as in M. abeloosi ) or with comparable length. For example, lengths measured on one side in two specimens for ungues I to IV are 14.9 µm, 14.8 µm, 18.1 µm, 20.3 µm and 15.9 µm, 17.0 µm, 18.1 µm, 20.2 µm, respectively.
Other traits (unless otherwise stated, elements reported here for M. grootaerti are lacking or not indicated in M. abeloosi ; frequency of some features not estimated): (1) bulb of femora III with adaxial, transverse and blunt carina located ventrally to porose area and distally to base of stalk, and touching ventrally border of ventroadaxial concavity of bulb or not (ventroadaxial concavity ordinarily well perceptible in M. grooaterti and also found in lectotype of M. abeloosi ; carina and concavity not shown in Fig. 3 View FIGURE 3 a); (2) genua I and II frequently with longitudinal and bent carina on abaxial side of stalk and reaching transverse furrow of bulb (see above) or not; (3) tibiae I and II with dorsal apparent outline of proximal region of bulb appearing flattened rather than concave as in M. abeloosi (according to Fig. 5 of Lions, 1978) in lateral orientation; (4) tibiae I usually with some (up to six on one side in one individual) dorsoproximal, transverse or oblique carinae on bulb (carinae with more or less long prolongation onto adaxial side are frequent whereas other carinae either have no prolongation or are prolonged onto abaxial side or onto both lateral sides); (5) bulb of tibiae II generally with one to three dorsoproximal, transverse or oblique carinae, and with distal, sinuous and transverse furrow made up of dorsal and convex distally part (on which solenidion φ is outwardly inserted) and of adaxial, convex proximally and less apparent part ordinarily passing proximally to anterolateral seta l’ and terminating slightly lower than l’; unlike M. abeloosi , no clear spot on adaxial side (Fig. 6c of Lions, 1978); (6) bulb of tibiae III usually with transverse furrow (located at some distance from proximal and transverse carina mentioned above, often on abaxial side or more seldom on both abaxial and ventral sides or possibly only on ventral side), and with longitudinal and roughly straight carina either dorsal (and then running to and ending near solenidion φ or not) or abaxial (and then extending to l’ and appearing to finish at some distance from l’); note that: (i) dorsal and abaxial carinae are occasionally observed together; (ii) according to some observations in SEM, carinae possibly come into contact with proximal and transversal carina of bulb; in lectotype of M. abeloosi , bulb of left tibia with proximoabaxial and longitudinal carina divided into two branches (dorsal branch running to solenidion φ and ventral branch terminating near l’); (7) proximal region of bulb in tibiae IV: dorsally, sometimes with transverse or oblique carinae (maximum five in one individual on one side) and infrequently with transverse furrow; abaxially or ventroabaxially, occasionally with small and roughly oblong concavity; (8) tarsi with furrows (for example, in tarsi I, transverse and dorsal furrow between solenidia and tectal setae (tc) and, in tarsi III, dorsoadaxial and longitudinal furrow proximally on bulb) and carinae (for example, in tarsi IV, one or two transverse and abaxial carinae dorsally to subunguinal seta s) not studied in detail; (9) in all legs, with guiding tube for flexor tendons ti of ambulacrum burrowed into ventral part of proximal border of tarsi.
Phaneres. Form of solenidia. Comparable in both species, except that in M. grootaerti , solenidia σ I and φ 2 are baculiform (instead of piliform and ceratiform, respectively, in M. abeloosi ; note that φ 2 are broadened over short distance proximally).
Labeling of setae: (1) seta labelled l’ in tarsi III by Lions (1978: Fig. 6a) might be anterior, fastigial seta ft’ having shifted in both ventral and proximal directions (see Grandjean, 1964b: Fig. 5a, for example in eupherederm Pheroliodes wehnckei ( Willmann, 1930) not closely related to genus Miracarus ); if hypothesis is correct, then no accessory seta is present in M. grootaerti and M. abeloosi ; (2) identification of seta l’’ in genua IV by Lions (1978: Fig. 6b) is wrong since seta l’’ is evidenced only in some macropyline Oribatida ( Grandjean, 1942a: 47, 51); accordingly, setae borne by genua IV in M. grootaerti as well as in lectotype of M. abeloosi are dorsal d and l’; note that presence of d and l’ in genua IV is common in Brachypylina whereas presence of d, l’ and v’ or of only d appears to be less frequent ( Grandjean, 1942a: 52).
Relative displacements (disjunctions and basculations) of pairs of setae (cf. Grandjean, 1958a). Disjunctions. Rules of disjunction for leg setae in Oribatida proposed by Grandjean (1960c: 273) are applicable to (v) in tibiae I– IV in both species (in tibiae II, somewhat less marked in M. grootaerti ; in tibiae III: less pronounced in M. grootaerti due to more distal insertion of v’’; distance between v’ and v’’: in M. grootaerti , mean = 4.4 µm, range: 3.4–5.9 µm, n = 5, and 8.5 µm in M. abeloosi ; in tibiae IV: strong disjunction in both species, and distance between v’ and v’’: in M. grootaerti , mean = 8.7 µm, range: 8.5–8.9 µm, n = 5, and 10.8 µm in M. abeloosi ) and appear to be not applicable in both species to following pairs of setae: (1) (l) in femora I–II (in femora I, double prime disjunction in M. grootaerti instead of either opposite, prime disjunction or lack of disjunction in M. abeloosi according to Figs 5a and 5c of Lions, 1978, respectively; note that disjunction is missing in both femora I in lectotype; in femora II, prime disjunction in both species; note that l’ and l’’ in femora II are characterized, besides, by clear displacement in dorsal direction); (2) (l) in genua I–II (in genua I, in M. grootaerti , very weak double prime disjunction or lack of disjunction as in M. abeloosi according to Fig. 5a of Lions, 1978; in genua II, weak or very weak double prime disjunction in M. grootaerti instead of poorly marked prime disjunction or outwardly obsolete disjunction in M. abeloosi according to Fig. 5b and 5e of Lions, 1978, respectively); (3) (l) in tibiae I–II (in tibiae I, in M. grootaerti , double prime disjunction very weak or disjunction absent as in M. abeloosi according to Fig. 5a of Lions, 1978; in tibiae II, double prime disjunction in both species); (4) (ft) in tarsi I–III (in tarsi I: in M. grootaerti , rarely double prime disjunction or more often either weak double prime disjunction or lack of disjunction; without disjunction in M. abeloosi according to Fig. 5a of Lions, 1978; in tarsi II, double prime disjunction seldom observed in M. grootaerti and not represented in Fig. 5b of Lions, 1978, in M. abeloosi ; in tarsi III, in both species, strong double prime disjunction at least if posteroabaxial seta is ft’ as indicated above); (5) primilateral setae (pl) in tarsi I (occasional and weak prime disjunction in M. grootaerti , and disjunction missing in M. abeloosi according to Fig. 5a of Lions, 1978; also detected in M. senensis according to Fig. 3 View FIGURE 3 a of Bernini, 1975); (6) primiventral setae (pv) in tarsi I–IV (double prime disjunction in tarsi I: infrequent and weak in M. grootaerti ; probably very weak in M. abeloosi , yet proximal part of pv’ is not distinctly shown in Fig. 5a of Lions, 1978; well detectable in M. senensis according to Fig. 3 View FIGURE 3 a of Bernini, 1975; in tarsi II, weak prime disjunction in M. grootaerti , and apparently disjunction absent in M. abeloosi according to Fig. 5b of Lions, 1978; in tarsi III, prime disjunction generally less marked in M. grootaerti ; in tarsi IV, ordinarily weak double prime disjunction in M. grootaerti instead of opposite, prime disjunction in M. abeloosi according to Fig. 6b of Lions, 1978.
Other pairs of setae: (1) (tc) in tarsi I–IV and (it) in tarsi I–III: in M. abeloosi , shifts hard to be identified in Figs 5 and 6 of Lions (1978); in M. grootaerti , shifts do not fit in with rule of disjunction since, in addition to being lacking, disjunctions appear to be very weak (and prime in (tc) and in (it) in tarsi I–II, and double prime in (tc) in tarsi III–IV), and infrequently weak and double prime in (tc) in tarsi III; (2) (p) in I–IV not studied in both species.
Disjunctions for which no rule is reported: (1) antelateral setae (a): prime disjunction in tarsi I–II in both species (and also in tarsi I in M. senensis according to Fig. 3 View FIGURE 3 a of Bernini, 1975; note that disjunction in tarsi II is rarely very weak in M. grootaerti ); in tarsi III, usually no disjunction in M. grootaerti and probably double prime weak disjunction in M. abeloosi according to Fig. 6a of Lions (1978); in tarsi IV, double prime disjunction very weak or disjunction missing in M. grootaerti and apparently double prime disjunction very weak in M. abeloosi according to Fig. 6b of Lions (1978); note that, in M. senensis , disjunction is prime in tarsi IV according to Fig. 3 View FIGURE 3 b of Bernini (1975); (2) ultimal setae (u): in M. grootaerti , no disjunction or very weak prime disjunction in tarsi I and II, and lack of disjunction in tarsi III and IV; in M. abeloosi , shifts not clearly shown in Figs 5 and 6 of Lions (1978) except weak prime disjunction represented in tarsi II in Fig. 5b.
Basculations. Prime basculations detected in: (1) (ft) in tarsi I (sometimes less pronounced in M. grootaerti ) and in tarsi II (rare and weak in M. grootaerti and outwardly very weak in M. abeloosi ); (2) (v) in tibiae IV (occasionally less marked in M. grootaerti ). Double prime basculations shown by: (1) (l) in femora I, in genua I and in tibiae I in both species; (2) (l) in genua II (in both species) and in tibiae II (sometimes less obvious in M. grootaerti ); (3) (v) in tibiae I (in both species) and in tibiae II (note that basculation is better marked in M. abeloosi since v’’ appears to be inserted beyond plane of pseudosymmetry, i.e. adaxially, in lateral projection, according to Fig. 5b of Lions, 1978, whereas v’’ being situated abaxially to plane of pseudosymmetry in M. grootaerti ); (4) (pl) in tarsi I (in both species and also in M. senensis according Fig. 3 View FIGURE 3 a of Bernini, 1975); (5) (pv) in tarsi II (in both species), in tarsi III (occasional and weak in M. grootaerti and apparently lacking in M. abeloosi according to Fig. 6a of Lions, 1978) and in tarsi IV (weak in M. grootaerti and not shown in Fig. 6b of Lions, 1978).
Other displacements of setae: (1) l’ and v’ in trochanters III apparently with variable placement in M. abeloosi since l’ and v’ appearing more distal and more proximal, respectively, in lectotype (as well as in M. grootaerti according to Fig. 3 View FIGURE 3 a) than in Fig. 6a of Lions (1978); note that (i) in M. grootaerti , variability of location of l’ is not negligible either (distance between proximal extremity of segment and seta/length of segment ratio ranges from 0.51 to 0.65, n = 4); (ii) in lectotype of M. abeloosi , position of l’ is appreciably different between both sides since ratio mentioned out above is 0.43 and 0.56 in right and left trochanter, respectively; (2) v’ in trochanters IV appearing more proximal in M. grootaerti in comparison with position shown in Fig. 6b of Lions (1978) and on both sides in lectotype of M. abeloosi ; (3) d in femora II shifted on abaxial side both in ventral and distal directions in M. grootaerti ( Fig. 3 View FIGURE 3 a) so that with placement clearly different from location in Fig. 5a of Lions (1978); (4) basifemoral seta ev’ in femora III more distal in M. grooaterti in comparison with Fig. 6a of Lions (1978) and lectotype of M. abeloosi ; (5) l’ in tibiae III usually with higher position in M. grootaeri than in Fig. 6a of Lions (1978) and on left side in lectotype of M. abeloosi ; (6) l’ in tibiae IV sometimes more dorsal in M. grootaerti in comparison with Fig. 6b of Lions (1978).
Doubling of setae: in trochanters III, v’ is doubled in one individual on one side and l’ is doubled in three other individuals on one side; in tibia IV, v’’ is doubled in one individual on one side.
Traits not studied in detail (yet succinct comparative survey conducted). Small depressions of apparent dorsal and ventral outline of segments (in lateral orientation) in connection with phaneres. Several are detected in both species like e.g. clear and regular depression of dorsal outline found in tarsi I in relation to solenidion ω 1, solenidion ω 2 and famulus e (see Fig. 5a of Lions, 1978, for M. abeloosi ). Another example of small depression of dorsal apparent outline is shown (yet partially) in Fig. 2 c referring to (tc) in tarsi IV in M. grootaerti (in M. abeloosi , depression appearing somewhat more distal according to Fig. 6b of Lions, 1978).
Size of phaneres. At rough estimate, length and thickness of phaneres comparable in both species (for example, solenidia σ IV are shortest solenidia whereas very long solenidia are only solenidia φ 1I, and l’ and l’’ in femora II and l’ in genua II are thickened) with few exceptions (e.g. l’’ in femora I and solenidia φ II appear shorter, and solenidia φ 1I and σ II appear thinner in M. grooaterti ).
Barbed setae. Barbs present in many setae in both species (according to comparison with lectotype of M. abeloosi , barbs omitted in several setae represented in Figs 5 and 6 of Lions, 1978); in M. grootaerti , a’ in tarsi I, III and IV, v’ in tibiae IV and pv’ in tarsi IV barbed (in contrast with M. abeloosi according to Lions, 1978: 417) whereas, in addition to eupathidic setae (p) and s in tarsi I and setae (p) in tarsi II, III and IV (see for example p’ in tarsus IV in Fig. 2 c), some setae such as d in genua IV and v’ in trochanters IV seem to be devoid of barbs; note that, in both species, barbs are often lined up (not clearly seen in setae of tarsus IV in Fig. 2 c).
Type material. The holotype and 10 paratypes, all females, were collected in grotto of ‘Vieux Banc’ in October 2006, and are deposited in the collections of the Royal Belgian Institute of Natural Sciences, Brussels (registration number: IG 31020).

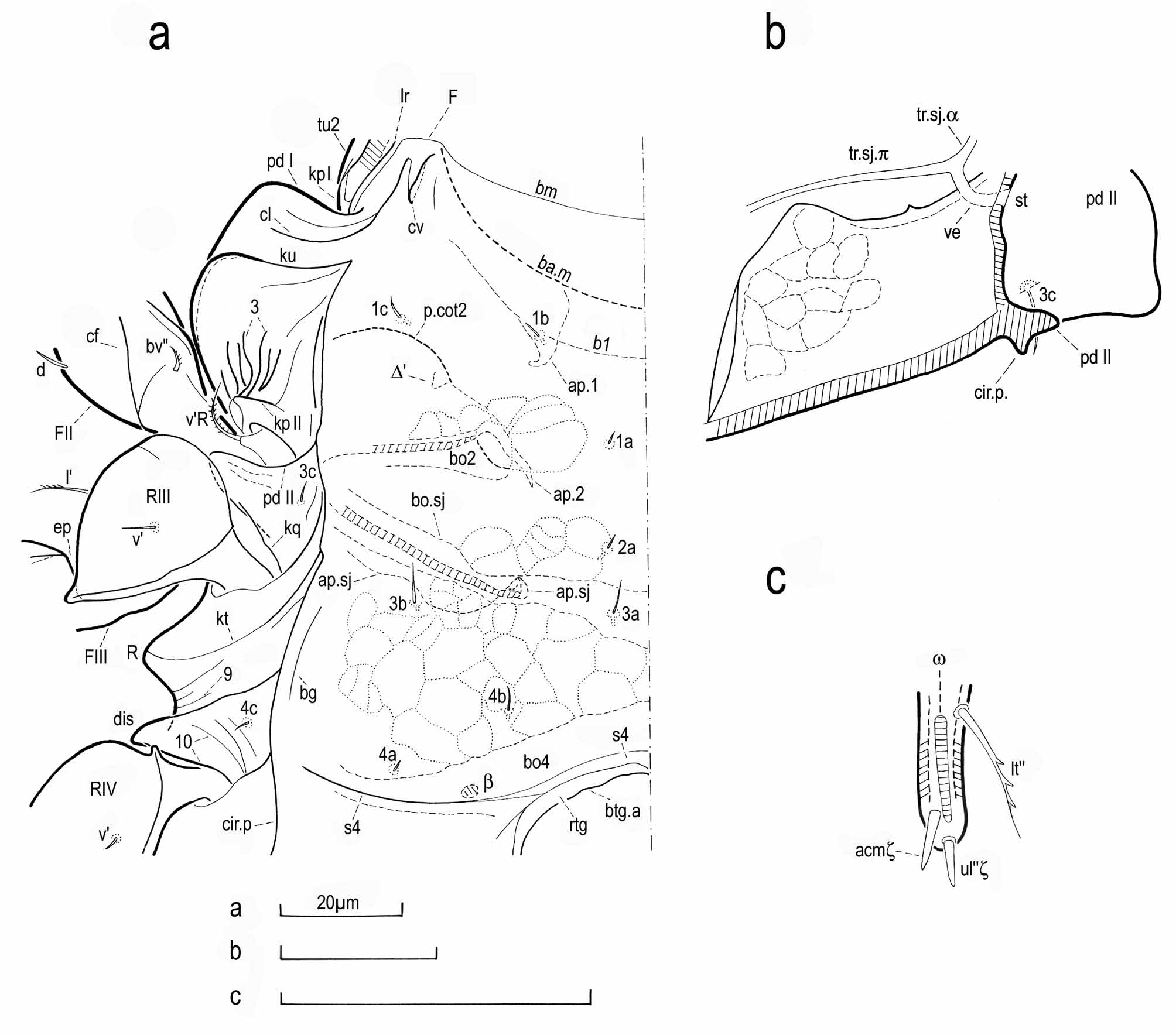

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.