
Limnodrilus profundicola ( Verrill, 1871 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4066.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:87790552-1DFF-41DF-B6C4-5BD43D19E998 |
|
DOI |
https://doi.org/10.5281/zenodo.3510454 |
|
persistent identifier |
https://treatment.plazi.org/id/03B77177-FFE5-A268-FF25-A916FE88F84C |
|
treatment provided by |
Plazi |
|
scientific name |
Limnodrilus profundicola ( Verrill, 1871 ) |
| status |
|
Limnodrilus profundicola ( Verrill, 1871)
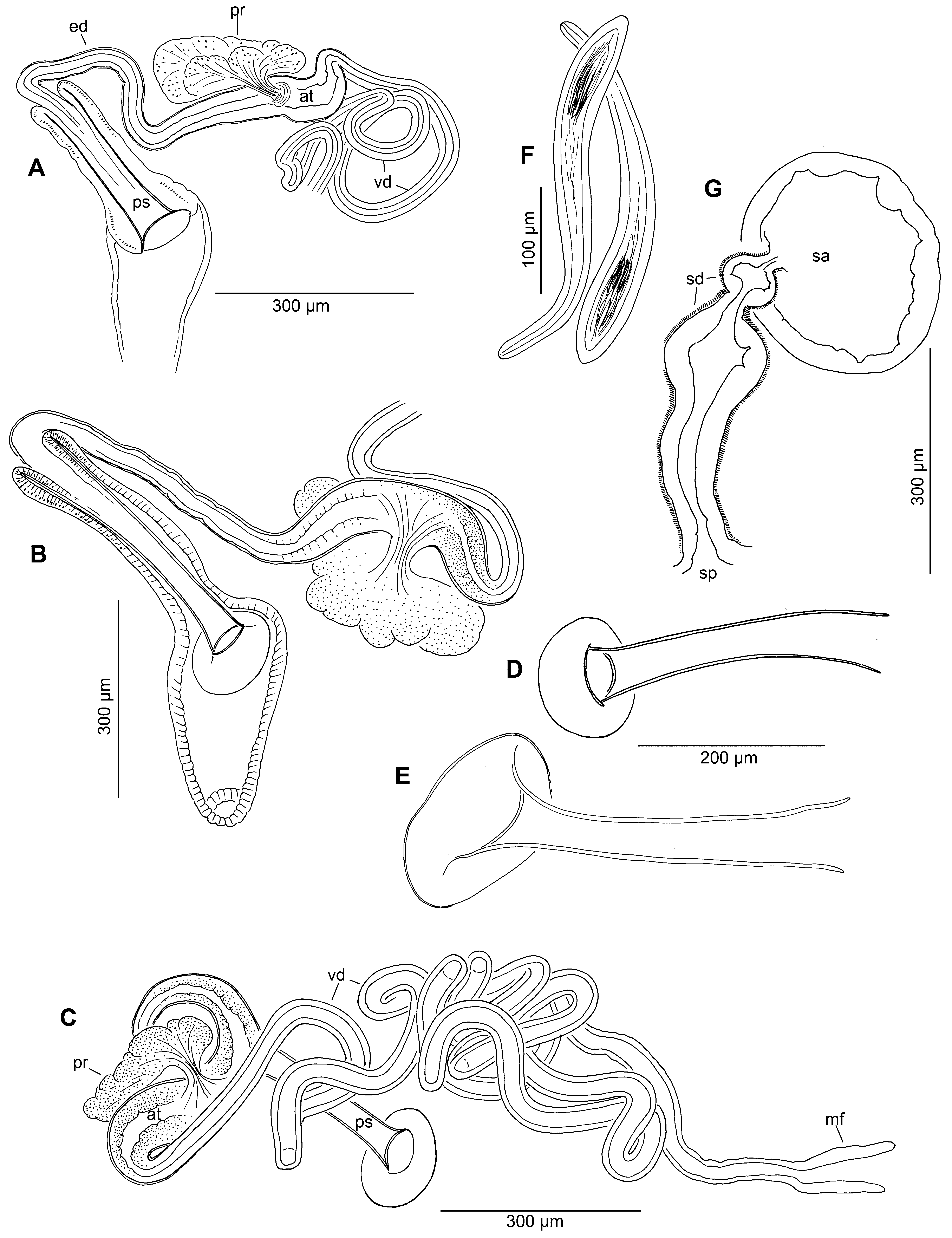
( Figure 4 View FIGURE 4 )
Tubifex profundicola Verrill, 1871 , in Smith & Verrill 1871: 451.
Limnodrilus helveticus Piguet, 1913: 134 –136, Fig. 8; Chekanovskaya 1962: 316 –317, Fig. 153A; Sokol’skaya 1972 (1983 translation): 11.
Limnodrilus profundicola (Verrill) . Brinkhurst 1965: 130 –131, Fig. 4 View FIGURE 4 K–M; Kennedy 1969: 54 –58, Figs. 5, 8, Tables 1 View TABLE 1 –3; Brinkhurst 1971: 470, Fig. 8.4D,E; Ohtaka 1992: 34 –35, Fig. 3 View FIGURE 3 , Steinlechner 1987: 57 –63, Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Timm 1998: 37, Figs. 68, 69; van Haaren & Soors 2013: 180 –181, Figs. 235–237; Cui et al. 2015: 526 View Cited Treatment –527, Fig. 7.
? Limnodrilus alpestris Eisen, 1879: 10 ; Eisen 1886: 896 –897, Pl. XII Fig. 11, Pl. XVII Fig. 11, Pl. XIX Fig. 18.
? Limnodrilus monticola: Eisen, 1879: 18 ; Eisen 1886: 896, Pl. XI, Fig. 10.
Material examined. Museum specimens. USNM 32598, Tubifex profundicola , holotype, 1 slide from Lake Superior. USNM 32597, Green Lake, Wisconsin, 1921, collected by Juday, whole mount, "referred to as? L. helveticus in notebooks made by F. Smith" ( Brinkhurst 1965). USNM 32590, Limnodrilus monticola , syntypes, 3 specimens from Sierra Nevada, California, 1876). USNM 32592, Limnodrilus alpestris , syntypes, 3 specimens from Sierra Nevada, California, 1876. All listed USNM material was examined by Brinkhurst (1965) and identified as Limnodrilus profundicola .
New material. USA, Lake Michigan, near Holland, Ottawa Co., Michigan, depth 6 to 25 m, Jun-1977, 2 whole mounts. Lake Michigan, nearshore habitat in the southern basin, various dates, collected by T. Nalepa, 6 dissected and 10 whole mounts. Estonia, Lake Peipsi, 6-Jun-1989, collected by T. Timm, 2 dissected, 3 whole mounts. Pärnu River, 28-Jul-1960, collected by T. Timm, 11 whole mounts. Russia, Arkhangelsk Region, Pinega River, 22-Jul- 1973, collected by T. Timm, 2 whole mounts. Kamchatka Peninsula, Lake Kuril’skoe, 27-Aug-1997, depth 3.5 m, collected by T.L. Vvdenskaya, 2 dissected, 1 whole mount. Japan, near Hyasaki, Lake Biwa, 90 m depth, 1-Dec- 1998, collected by. A. Ohtaka, 2 dissected, 2 whole mounts.
Descriptive notes. Museum specimens: The holotype, USNM 32598, has mostly dissolved since Brinkhurst’s (1965) redescription, with no penis sheath or chaetae visible. USNM 32597 probably represents a different species, as the penis sheaths are about 500 mm long, curved, with length/width ratio over 8. USNM 32590 and 32592 could not be confirmed, as penis sheaths were not visible.
New material: Chaetae are bifid with teeth slightly curved and approximately equal, or the upper tooth slightly longer in anterior segments ( Fig. 4 View FIGURE 4 G–J). As in most naidid species, chaetae are more numerous and slightly larger in bundles of anterior segments, and there are at most 1 or 2 "replacement" chaetae. Number of chaetae per bundle 4–9 in anterior segments, 2–3 in posterior segments.
Chloragogen tissue is not particularly dark in any of the profundicola specimens. It densely covers the gut beginning in VI, although it may be visible dorsally in V.
Length and proportions of penis sheaths are consistent with published values ( Fig. 4 View FIGURE 4 A–D, Table 1 View TABLE 1 ). The head is always round and symmetrical, well-defined and orthogonal to the straight or slightly curved shaft, and usually somewhat curved back ( Fig. 4 View FIGURE 4 A–D,K). Musculature is weak or not apparent in the penial sac, and does not show the strong spiral pattern seen in some other Limnodrilus species. Atria appear relatively short for the genus ( Fig. 4 View FIGURE 4 A–D, Table 1 View TABLE 1 ).
The spermathecal ducts appear quite variable in general form. Still, the structure consists of an irregular sac covered with a thick layer of mostly circular muscles, which narrows to a variably expanded duct before joining the ampulla ( Fig. 4 View FIGURE 4 E,F). The irregular ectal part ranges from globose to narrow within populations, and in some specimens it is similar to the corresponding part in the L. sulphurensis material (cf. Fig. 2 View FIGURE 2 G).
Remarks: The original L. profundicola description (Verrill, in Smith & Verrill 1871), based on material from Lake Superior, gives no details on reproductive characters used to distinguish Limnodrilus species. As the holotype and most other museum specimens examined no longer appear useful, diagnostic characters given in more recent accounts are difficult to verify independently. Furthermore, measurements given in Brinkhurst’s (1965) North American key and 1971 redescription may reflect those derived from Britain and continental Europe ( Kennedy 1969), rather than topotypic or even Nearctic populations. The characters given by Brinkhurst (1971) refer to the form of the chaetae ("upper tooth at most a little longer than lower") and then to the proportions of the penis sheaths. Approximate measurements of penis sheaths in Fig. 4 View FIGURE 4 K–M in Brinkhurst (1965) indicate total lengths about 270 µm (for profundicola , presumably based on the holotype), 340 µm for profundicola (as monticola ), and 240 µm for profundicola (as helveticus , presumably European). The range of penis sheath length/width values is very broad (2–7) in those references, but the mean (4.5) is similar to other published profundicola values, as well as to the new measurements ( Table 1 View TABLE 1 ). Kennedy (1969) appears to use the same values, concluding that profundicola is best determined by the circular head at the ectal end of the sheath, which is more broad and flat than that of L. udekemianus , and never forming an asymmetrical hood, as in some forms of L. hoffmeisteri sensu lato.
The description of L. helveticus ( Piguet 1913) was more detailed, showing basic morphology of the penis sheath, atria, chaetae, and spermathecae. It should be noted that Sokol’skaya (1972, 1983) questioned Brinkhurst’s synonymies and preferred to retain helveticus , based on the high variability in penial morphology in the above descriptions, and also because the profundicola spermathecae were not described by either Brinkhurst (1965, 1971) or Kennedy (1969). This apparently refers to the spermathecal duct, which Sokol’skaya (1983, Fig. 6) describes and illustrates as "forming a distal expansion". Piguet (1913, Fig. 8B) described and illustrated a duct with a globular ectal end, and Cui et al. (2015, Fig. 7A) illustrate a sharply expanded spermathecal duct in a specimen from Tibet, attributed to profundicola . Spermathecal ducts were consistently enlarged in all of our new material, but the ectal end was globular in some, sacciform in other specimens ( Fig. 4 View FIGURE 4 E,F).
The new Nearctic material is from southern Lake Michigan, rather than the type locality in Lake Superior, but still within the Great Lakes region. Size and proportions of the penis sheath and chaetal morphology were reasonably consistent among regions, and comparable to literature values ( Table 1 View TABLE 1 ), thus supporting Brinkhurst’s (1966) combination of helveticus with profundicola . Penis sheaths of the new material are all similar to illustrations by Brinkhurst (1965, Fig. 4 View FIGURE 4 K–M), Piguet (1913, Fig. 10b), van Haaren & Soors (2013, Fig. 237) and Cui et al. (2015, Fig. 7B). Chaetae are typical for the genus; teeth have normal, slight curvature, in contrast to the strongly angled teeth in L. sulphurensis . The nearly equal teeth in new specimens ( Fig. 4 View FIGURE 4 G–J) generally resemble Fig. 3 View FIGURE 3 A–B of Ohtaka (1992), Fig. 68 in Timm (1998) and Fig. 4 View FIGURE 4 in Steinlechner (1987), but contrast slightly with Fig. 8a of Piguet (1913) and Fig. 5 of Kennedy (1969), in which the upper tooth is distinctly longer. The chaetae in anterior bundles show a fan-like orientation, considered characteristic of Palearctic profundicola populations ( Fig. 4 View FIGURE 4 G; T. Timm, personal communication). The spermathecal duct is expanded ( Fig. 4 View FIGURE 4 F), consistent with Palearctic populations, and further supporting their combination.


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Tubificinae |
|
Genus |
Limnodrilus profundicola ( Verrill, 1871 )
| Fend, Steven V., Liu, Yingkui, Steinmann, David, Giere, Olav, Barton, Hazel A., Luiszer, Fred & Erséus, Christer 2016 |
Limnodrilus profundicola
| Cui 2015: 526 |
| Haaren 2013: 180 |
| Timm 1998: 37 |
| Ohtaka 1992: 34 |
| Steinlechner 1987: 57 |
| Brinkhurst 1971: 470 |
| Kennedy 1969: 54 |
| Brinkhurst 1965: 130 |
Limnodrilus helveticus
| Chekanovskaya 1962: 316 |
| Piguet 1913: 134 |
Limnodrilus alpestris
| Eisen 1886: 896 |
| Eisen 1879: 10 |
Limnodrilus monticola:
| Eisen 1886: 896 |
| Eisen 1879: 18 |
Tubifex profundicola
| Smith 1871: 451 |