
Neverita duplicata, (SAY 1822)
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00058.x |
|
DOI |
https://doi.org/10.5281/zenodo.5490934 |
|
persistent identifier |
https://treatment.plazi.org/id/03B6B923-EE20-FFFA-8CD0-63853F976BE3 |
|
treatment provided by |
Carolina |
|
scientific name |
Neverita duplicata |
| status |
|
NEVERITA DUPLICATA (SAY 1822) View in CoL
Material examined
Wood’s Hole, Massachusetts ( USNM 890941). Only immature males and females were available for study. Thus, observations on reproductive anatomy are preliminary.
External anatomy and mantle cavity
Mantle edge smooth with single, small pallial tentacle lying within exhalent margin. Hypobranchial gland weakly folded. Bipectinate osphradium lying near inhalant margin, approximately one-half length of ctenidium. Foot massive with extensive propodium. Anterior edge of propodium sharply indented at midline. Anterior pedal gland opening along junction of propodium and metapodium. Large, horny operculum present. Cephalic tentacles flattened and triangular with shallow, ventral groove extending along outer edges. Two broad, flattened lobes present on either side of cephalic tentacles, bearing recessed eyes along ventral surfaces. Eyes weakly pigmented, retaining lens. Rhynchostome opening ventrally.
Reproductive system
Gonad dorsally overlying digestive gland on right. Vas deferens forming convoluted seminal vesicle. No gonopericardial connection present in females or males. Glandular pallial gonoduct open in both sexes. No sperm storage structures apparent associated with pallial or renal oviduct. Closed pallial vas deferens crossing neck from open prostate to penis behind right cephalic tentacle.
Alimentary system
Foregut. Radula taenioglossate. Foregut modified into short acrembolic proboscis lying over circum-oesophageal nerve ring when retracted. Transverse mouth opening at tip of extended proboscis, surrounded by tumid outer lips with ventral cleft. Accessory boring organ present under lip, forming large flattened pad bearing rodlet border. Large, paired jaws present at distal tips of dorsal folds. Sublingual cavity shallow. Subradular membrane incompletely covering surface of odontophore. Small subradular organ present ( Figs 4F View Figure 4 , 5G View Figure 5 , sro), forming rounded, folded and glandular protuberance. Anterior oesophagus flattened, bearing ciliated dorsal folds and dorsal food groove and non-ciliated ventral channel ( Fig. 10A View Figure 10 , vc); no ventral folds present. Two small, ciliated ducts emerging from ascinous salivary gland, passing through nerve ring embedded in dorso-lateral walls of oesophagus. Large odontophoral retractor muscle attaching ventrally to buccal mass and splitting, inserting onto lateral walls of cephalic haemocoel. Large, septate oesophageal gland present within mid-oesophagus. Surface of septae sculptured with fine, anastomosing channels. Gland differentiated into small anterior light coloured region and posterior, brown region. Dorsal food groove, u-shaped and enclosed by one of flanking folds.
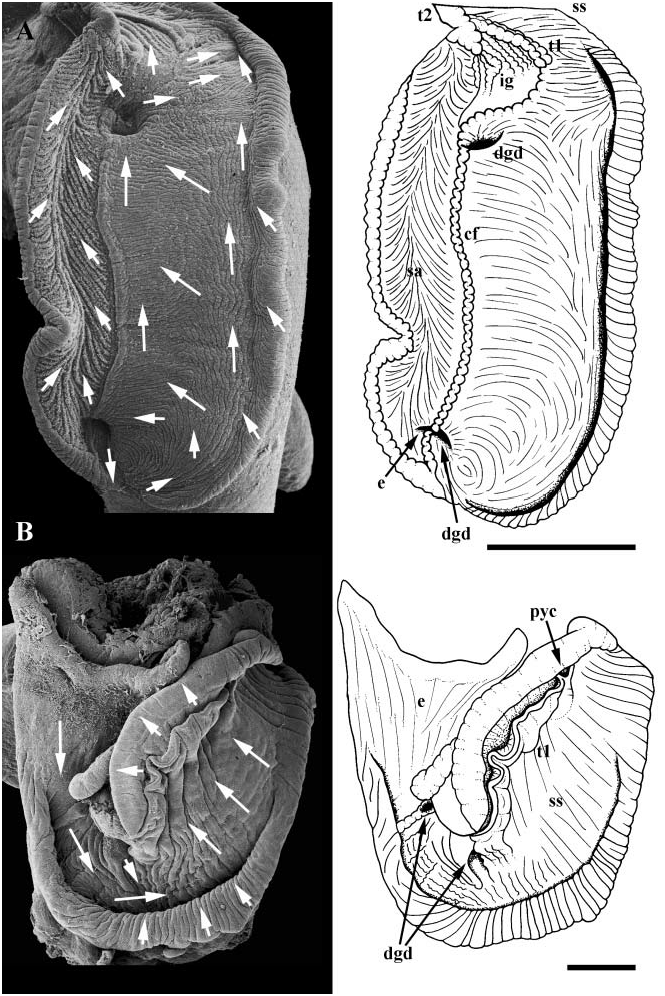
Midgut. Oesophagus ( Fig. 16A, e View Figure 16 ) entering posterior end of narrow, elongate midgut. Gastric shield absent. Left, ventral wall of gastric chamber lined with welldeveloped sorting area (sa), extending from oesophageal aperture to intestinal groove (ig). Sorting area flanked on right by single, low ciliated fold (cf). Ciliated fold extending between posterior digestive gland duct (dgd), opposite oesophageal aperture, and anterior duct near minor typhlosole (t2). Sorting lamellae comprising two sets of parallel folds, directed anteriorly and obliquely toward midline of sorting area. Ciliary currents within sorting area flowing toward intestinal groove. Currents within gastric chamber flowing anteriorly along right wall and clockwise along ventral floor. Style sac region (ss) comprising two, low typhlosoles (t1, t2) bounding short, narrow region of transverse folds bearing low, compact cilia. Ciliary tract on major typhlosole absent. Currents flowing clockwise within style sac region, posteriorly within intestinal groove.
Hindgut. Intestine emerging from style sac, curving slightly posteriorly and dorsally along rear wall of kidney and pericardium, then continuing anteriorly.
Reno-pericardial system
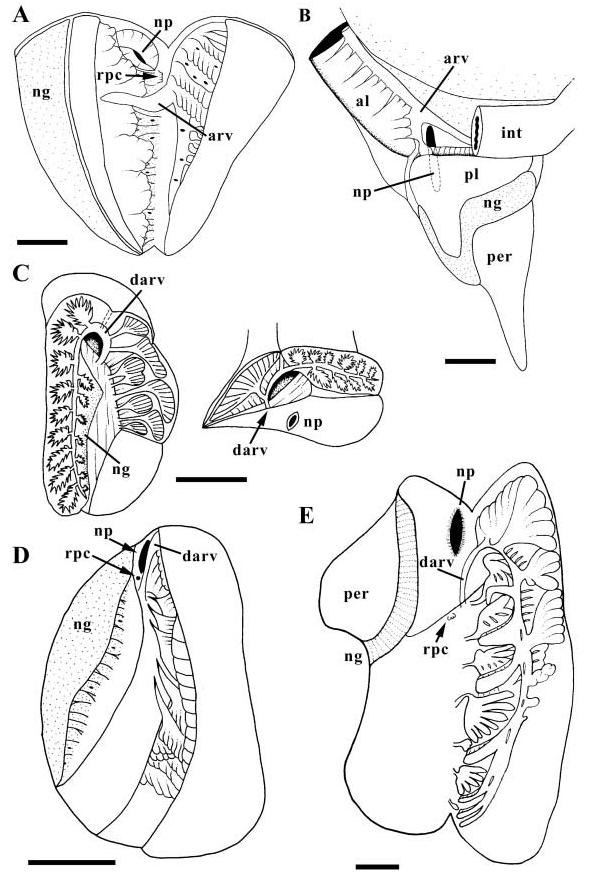
Excretory tissue of kidney divisible into three parallel, histologically distinct lobes ( Fig. 23A View Figure 23 ). Right lobe dorsally surrounding intestine, comprising folded, tubular excretory lamellae. Central lobe anteriorly overhanging nephropore, comprising villous, massive excretory lamellae. Lobe bordering pericardium on left side of kidney, comprising hypertrophied nephridial gland (ng). Afferent renal vessel entering kidney floor anteriorly, just behind reno-pericardial canal (rpc). Afferent renal vessel splitting into two large branches, one extending antero-dorsally along front wall of kidney and supplying right lobe, second branch extending postero-dorsally. Postero-dorsal branch supplying both right and central lobes.
Nervous system and sensory structures
Nervous system epiathroid, left zygoneurous. Nerve bundle emerging from each cerebral ganglion containing tentacular and optic nerves. Tentacular and optic nerves becoming distinct short distance from nerve ring. Single tentacular nerve producing numerous, small branches innervating tentacle and cephalic haemocoel. Four nerves from each cephalic ganglion innervating proboscis; one representing long buccal connective. Large pedal ganglia adjacent to cerebral and pleural ganglia; pedal connectives short. Prominent propodial ganglia present at anterior ends of pedal ganglia. Pedal nerves ganglionated. Sub-oesophageal and pleural ganglia adjacent; supra-oesophageal connective long. Pallial nerves from sub-oesophageal and right pleural ganglia contacting, but no dialyneury formed. Visceral loop bearing two asymmetrical visceral ganglia straddling posterior oesophagus. Statocysts with single statoliths lying ventrally and anterior to circum-oesophageal nerve ring.
Discussion
Much of the literature pertaining to naticids concerns their predatory behaviour. Accordingly, anatomical descriptions are largely restricted to foregut anatomy – the radula and accessory boring organ in particular. Thus, few comprehensive anatomical studies are available, with only rare accounts of reproductive, renal, and nerve morphology.
The naticid foregut is characterized by the presence of an acrembolic proboscis, large jaws and an accessory boring organ ( Hirsch, 1915; Eales, 1923; Ziegelmeier, 1954; Fretter & Graham, 1962; Marcus & Marcus, 1965). The organ has been found to consist of two distinct epithelia; one secretory and one comprising a microvillous brush border extending through a matrix of protein and chitin ( Bernard & Bagshaw, 1969). This and other studies have found a common fine-structural construction between the proboscidial accessory boring organ of naticids and the pedal accessory boring organ of muricids ( Carriker, 1981). Glands associated with the foregut include an ascinous salivary gland and a large oesophageal gland ( Bouvier, 1887; Hirsch, 1915; Eales, 1923; Ziegelmeier, 1954; Fretter & Graham, 1962; Marcus & Marcus, 1965; Reid & Friesen, 1980). The oesophageal gland is differentiated into two discrete regions and comprises septae which are finely sculptured with radiating canals (Reid & Friesen). The dorsal food groove is ushaped (Fretter & Graham; Reid & Friesen).
The midgut of naticids is elongate with the oesophageal aperture opening at the posterior tip; a gastric shield is absent ( Simroth, 1896 –1907; Hirsch, 1915; Fretter & Graham, 1962; Reid & Friesen, 1980). Several aspects of midugt morphology vary within the family. The large sorting area may extend the full length of the gastric chamber along the left (Reid & Friesen), or may be restricted to a rather small area near the intestinal groove (Fretter & Graham). The digestive gland ducts may open at opposite ends of the sorting area (Hirsch; Reid & Friesen), or both may open near the intestinal groove (Fretter & Graham). The ciliated fold to the right of the sorting area is accompanied by a groove that joins the midline rejection tract of the sorting area (Reid & Friesen); no distinct groove is present in Neverita duplicata . Fretter & Graham reported that only vestiges of the typhlosoles and style sac region remain. Similarly, Reid & Friesen reported that no trace of a style sac region is apparent, but clearly show ciliary currents flowing in a clockwise direction in this portion of the gut.
The excretory tissue is subdivided into two lobes that are histologically and macroscopically distinct ( Simroth, 1896 –1907) – the so-called folded and absorbing kidney lobes of Fretter & Graham (1962). A nephridial gland is present; although extremely large in N. duplicata , the gland may be restricted to a relatively narrow band along the pericardial wall (Fretter & Graham). The afferent renal vessel splits to supply both lobes (Fretter & Graham).
The naticid nervous system is epiathroid and left zygoneurous with long buccal connectives ( Bouvier, 1887; Eales, 1923). Three cerebral nerves innervate the proboscis (Bouvier). Sensory structures include a large bipectinate osphradium with leaflets of a complex structure ( Marcus & Marcus, 1965; Maeda, 1988, 1990). Eyes have been reported as lacking ( Marcus & Marcus, 1965), although this may be a consequence of the fact that they are recessed within flattened lobes at the outer bases of the cephalic tentacles (present study). Similar lobes have been recorded in several species (e.g. Ziegelmeier, 1957).



| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.